
Reciprocal Analysis of Infections of a Model Reveal Host-Pathogen Conflicts Mediated by Reactive Oxygen and imd-Regulated Innate Immune Response
The survival of a bacterial pathogen within a host depends upon its ability to outmaneuver the host immune response. Thus, mutant pathogens provide a useful tool for dissecting host-pathogen relationships, as the strategies the microbe has evolved to counteract immunity reveal a host's immune mechanisms. In this study, we examined the pathogen Francisella novicida and identified new bacterial virulence factors that interact with different parts of the Drosophila melanogaster innate immune system. We performed a genome-wide screen to identify F. novicida genes required for growth and survival within the fly and identified a set of 149 negatively selected mutants. Among these, we identified a class of genes including the transcription factor oxyR, and the DNA repair proteins uvrB, recB, and ruvC that help F. novicida resist oxidative stress. We determined that these bacterial genes are virulence factors that allow F. novicida to counteract the fly melanization immune response. We then performed a second in vivo screen to identify an additional subset of bacterial genes that interact specifically with the imd signaling pathway. Most of these mutants have decreased resistance to the antimicrobial peptide polymyxin B. Characterization of a mutation in the putative transglutaminase FTN_0869 produced a curious result that could not easily be explained using known Drosophila immune responses. By using an unbiased genetic screen, these studies provide a new view of the Drosophila immune response from the perspective of a pathogen. We show that two branches of the fly's immunity are important for fighting F. novicida infections in a model host: melanization and an imd-regulated immune response, and identify bacterial genes that specifically counteract these host responses. Our work suggests that there may be more to learn about the fly immune system, as not all of the phenotypes we observe can be readily explained by its interactions with known immune responses.
Published in the journal:
. PLoS Pathog 6(8): e32767. doi:10.1371/journal.ppat.1001065
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1001065
Summary
The survival of a bacterial pathogen within a host depends upon its ability to outmaneuver the host immune response. Thus, mutant pathogens provide a useful tool for dissecting host-pathogen relationships, as the strategies the microbe has evolved to counteract immunity reveal a host's immune mechanisms. In this study, we examined the pathogen Francisella novicida and identified new bacterial virulence factors that interact with different parts of the Drosophila melanogaster innate immune system. We performed a genome-wide screen to identify F. novicida genes required for growth and survival within the fly and identified a set of 149 negatively selected mutants. Among these, we identified a class of genes including the transcription factor oxyR, and the DNA repair proteins uvrB, recB, and ruvC that help F. novicida resist oxidative stress. We determined that these bacterial genes are virulence factors that allow F. novicida to counteract the fly melanization immune response. We then performed a second in vivo screen to identify an additional subset of bacterial genes that interact specifically with the imd signaling pathway. Most of these mutants have decreased resistance to the antimicrobial peptide polymyxin B. Characterization of a mutation in the putative transglutaminase FTN_0869 produced a curious result that could not easily be explained using known Drosophila immune responses. By using an unbiased genetic screen, these studies provide a new view of the Drosophila immune response from the perspective of a pathogen. We show that two branches of the fly's immunity are important for fighting F. novicida infections in a model host: melanization and an imd-regulated immune response, and identify bacterial genes that specifically counteract these host responses. Our work suggests that there may be more to learn about the fly immune system, as not all of the phenotypes we observe can be readily explained by its interactions with known immune responses.
Introduction
The outcome of any infection, whether it be clearance of the infecting pathogen, establishment of a persistent infection, or even death of the host is determined by contributions from both the host and the microbe [1]. To infect a susceptible host microbes express virulence factors, genes that allow the pathogen to invade, colonize, and survive within the host and cause essential pathology. In response, the host initiates an immune response that attempts to clear the pathogen and increase tolerance to the ensuing infection [2]. Consequently, in addition to genes that allow the bacteria to invade host cells and obtain nutrients from its host, a subset of the virulence factors expressed by the microbe must address the need of the bacteria to counteract the host immune response. Exploring this complex interplay between host and pathogen can help us to understand bacterial pathogenesis and define the contributions of the host immune system to bacterial virulence.
One way to explore the host-pathogen relationship is to apply model systems that allow us to dissect the genetics of both sides of the equation simultaneously in vivo [3]. In this study, we examine the host-pathogen interactions of Francisella novicida with an insect host, Drosophila melanogaster, and identify aspects of fly immunity that are most important for fighting F. novicida infection as well as the bacterial virulence factors that interact with each of these specific immune responses. Drosophila is used as a model of innate immunity because its simplicity and the ease at which it can be used for both forward and reverse genetics has allowed for the identification and characterization of aspects of the innate immune response that are conserved across evolution [4]–[6]. The fly immune response has three effector arms: an inducible antimicrobial peptide (AMP) response, a reactive oxygen response mediated by the activation of the enzyme phenoloxidase and the deposition of the pigment melanin, and a cellular immune response in which foreign invaders are phagocytosed by Drosophila hemocytes [7]. The humoral AMP response has been studied extensively and shown to be regulated by two pattern recognition pathways, Toll and imd which have been well-characterized and described, but the regulatory mechanisms of the melanization and cellular immune responses have only recently become the focus of increased interest and have not yet been fully elucidated [8].
Previous studies with pathogenic bacteria in the fly have shown that virulence factors that function in the vertebrate hosts of these pathogens are often required for the microbe to survive in the fly [6]. Recently, this has been shown to be true for the live vaccine strain (LVS) of the virulent pathogen Francisella tularensis, a Gram-negative facultative intracellular bacterium that is the causative agent of tularemia [9]. F. tularensis can infect a wide range of hosts that includes humans, but is more commonly associated with rabbits and small rodents. Unlike many of the pathogens used in previous fly studies, F. tularensis also has a documented arthropod vector phase in its natural life-cycle [10], [11]. While many arthropod-vectored pathogens can only be transmitted by a single specific species, F. tularensis is able to infect arthropods ranging from ticks to multiple species of mosquito to biting flies such as deerflies [12]–[14]. This makes the Drosophila model system particularly useful for studying both general F. tularensis host-pathogen interactions and insect-specific factors.
To date, the fly has primarily been used to dissect the function of known bacterial virulence factors or to demonstrate conservation between fly and vertebrate defenses. Less has been done to use forward genetic approaches in the microbe to identify virulence factors de novo. As immunologists we tend concentrate on known signaling pathways that have proven simple to study, are of interest to those working in vertebrates because they are conserved, and those that fit our idea of what the fly's immune response should be. In other words, experiments are driven by the interests of the scientists and not the pathogens. We took a more ecology-based approach and determined, from scratch, what this fly pathogen needs to kill the fly. The advantage of the fly is that it is inexpensive, rapid to use and has extensive genetic tools. The fly could be a useful tool for the identification of new virulence factors rather than a system used to study known factors.
To identify new virulence factors and examine their interactions with the fly immune system, we used the Francisella novicida strain U112 to infect flies and performed a genome-wide screen to identify factors required for growth and/or survival within the fly. Many of the genes that we identified are required for resistance to the Drosophila innate immune response, particularly to the oxidative stress produced by melanization. This is interesting in particular, because until recently, this pathway had been discarded as having no relevance to microbial infections in the fly [15]–[17]. Our work demonstrates that bacterial mutants can be used as probes of the host immune system to identify what aspects of innate immunity are most important in determining the outcome of an infection.
To identify additional interactions between the host immune system and bacterial virulence factors, we performed a second genetic screen in which we compared the ability of F. novicida mutants to grow in wild type flies to flies with an immunity defect known to affect fly survival in F. tularensis infections. These flies lack a functional imd signaling pathway, and we anticipated that this would reveal bacterial mechanisms necessary to circumvent the imd-regulated immune response. The imd pathway is primarily described as inducing antimicrobial peptides. Although we identified bacterial genes required to resist antimicrobial peptide killing in vitro, we were particularly intrigued to find a subset of genes that when mutated did not appear to change F. novicida sensitivity to the antimicrobial peptide we tested yet showed an altered phenotype in imd mutants. This suggests the possibility that there are previously undescribed immune mechanisms that are regulated by the imd pathway.
Results
Characterization of Francisella tularensis ssp. novicida infections of Drosophila melanogaster
We infected flies with F. novicida strain U112, a wild type strain that causes virulent infections in its natural mouse and rabbit hosts but is not pathogenic to humans. Previous work using the Live Vaccine Strain (LVS) of F. tularensis demonstrated that F. tularensis grows to high bacterial levels within flies and causes a lethal infection [9]. Infections of the fly with the U112 strain were consistent with this result, although we found the U112 strain to be slightly more virulent in Drosophila than the LVS strain, killing the fly with a median time to death (MTD) of 5 days post-infection with 103 CFU (Figure 1A and Figure S1). As few as 5 CFU of F. novicida U112 were sufficient to kill the fly, and bacterial growth within the fly was exponential approaching 5×107 CFU per fly before they succumbed to the infection. Regardless of the initial dose, F. novicida infections quickly reached the same high levels of bacteria; colony counts in flies receiving a low dose caught up to the 108 fold higher dose within two days (Figure 1B and Figure S2).

In mammalian infections, F. tularensis is a facultative intracellular parasite that primarily grows within macrophages [18]. However, due to the extremely high bacterial levels observed within the fly we speculated that a large proportion of the bacteria were growing extracellularly. To test this idea, we performed gentamycin chase assays, infecting flies with F. novicida and subsequently injecting them with the non-cell permeable antibiotic gentamycin at various timepoints post infection. This assay kills off extracellular bacteria while leaving bacteria growing within cells intact and allowing us to determine whether the bacteria are growing intracellularly or extracellularly By 24 hours post infection, approximately 1×104 CFU per fly were found to be localized intracellularly. However, the average total population of bacteria in infected flies was at least 1×105 CFU per fly, demonstrating that a significant population of bacteria was located extracellularly. Over the course of the infection the total bacteria population increased to 1×107 CFU/fly, while the intracellular population remained steady, indicating that the extracellular population was responsible for the exponential increase in bacteria seen during fly infections (Figure 1C). This is roughly similar to what has been observed using the LVS strain, although the absolute numbers differ slightly possibly due to the differences in virulence between the two bacterial strains, differences in the host strains or environmental conditions [9].
Identification of F. novicida virulence factors with a Transposon Site Hybridization (TraSH) screen
Having demonstrated that the fly can support F. novicida growth we applied this model to the identification of bacterial genes that were important for establishing infection within the fly. Previous work has shown that many F. tularensis virulence factors that are required for growth in mammalian models are also essential in insect infections, including the Francisella Pathogenicity Island (FPI) genes iglB, iglC, and iglD and the transcription factor mglA [9], [19]. To expand upon this work, we sought to identify additional bacterial virulence factors and provide an opportunity to discover new biology using a forward genetic approach.
Using a transposon insertion library of F. novicida mutants we performed an in vivo screen for mutants with altered growth rates compared to wild-type bacteria using a Transposon Site Hybridization (TraSH) assay. Briefly, flies were infected with the pooled library and the infection was allowed to proceed for two days, at which point the bacterial populations in each fly were harvested. Genomic DNA was then purified from this population of bacteria and from the original input library that had not been subjected to the stresses found within the fly. RNA was amplified from the site of each transposon insertion and the two populations of RNAs were compared by microarray analysis. We identified mutants representing 149 F. novicida genes that were negatively selected with a false discovery rate (FDR) of 5%, indicating that these genes were essential for bacterial growth and survival within the fly (Table S1). 41 of these genes had previously been identified as negatively selected in a similar TraSH analysis performed with the same mutant library in an in vivo mouse model; this list includes the known virulence genes iglC, iglD, pdpA, and pdpB, and mglA [20]. In addition, 11 genes from our screen overlapped with data from a negative selection screen performed by Kraemer et al. using an inhalation model to observe respiratory infections in the mouse, and an additional 8 overlapped with a signature-tagged mutagenesis screen performed by Su et al. that also used an intranasal route of inoculation [21], [22]. The overlap between our TraSH assay and additional Francisella genome-wide screens is shown in Table 1 and Figure 2. Interestingly, no genes were identified in all four unique screens, although 7 genes were identified in our fly TraSH and at least two other screens. These genes were the hypothetical proteins FTN_1682 and FTN_1016, the RNA methyltransferase yibK, the ABC transporter yjjK, the amino acid antiporter FTN_0848, iglC and iglD. The degree of overlap between our fly screen and similar mouse screens both supports the hypothesis that our screen in Drosophila can be used to identify virulence factors that are conserved between insect and mammalian infections, and also presents the possibility of identifying virulence factors unique to the arthropod vector stage of the F. novicida life cycle.


Confirmation of negatively selected mutants
To confirm the results of the TraSH screen, we tested 65 of the negatively selected mutants individually by competition assay, focusing on mutants that had particularly large decreases in abundance and/or showed homology to bacterial genes that we predicted could play a role in immune evasion or modulation. Transposon insertion mutants of each gene containing a kanamycin resistance cassette were tested to determine their ability to grow in competition with wild-type F. novicida. Each mutant was mixed with wild-type bacteria and injected into wild-type Drosophila at a 1∶1 ratio. Infected flies were incubated for 48 hours, at which point the bacteria in each fly were plated and the ratio of mutant to wild-type bacterial was determined. A competitive index of 1 represents an equal ratio of mutant to wild-type bacteria, while competitive indexes of less than one indicate that the mutant is attenuated. Mutants that were determined to be statistically significantly less than 1 by one sample t-test in a minimum of three repetitions were considered attenuated and are listed in Figure 2.
Mutants that were confirmed as negatively selected included kdpA, kdpC, kdpD, and kdpE, components of 2-component regulatory system that responds to turgor pressure, a number of genes known to be regulated by the virulence factor mglA, members of the Major Facilitator Superfamily (MFS) thought to be involved in substrate transport and drug resistance, multiple genes know to be involved in DNA repair, and a number of hypothetical proteins. (Figure 3A and data not shown) 56 of the mutants tested showed attenuated phenotypes by competition assay, with competitive indexes ranging from 0.6-0.007. The results of the competition assays indicated that the microarray data produced by the TraSH assays is useful for predicting negatively selected mutants but was somewhat non-quantitative; the degree of attenuation as measured by microarray analysis did not always correlate with the strength of the phenotype observed by competition assay.

F. novicida mutants demonstrating increased sensitivity to oxidative stress are attenuated in Drosophila
One set of negatively selected mutants stood out as particularly interesting because they indicated a bacterial requirement for resistance to oxidative stress within the fly. These mutants included the LysR family transcriptional regulator oxyR. The homologue of this gene in E. coli has been shown to sense hydrogen peroxide and induce the transcription of downstream genes that provide protection against oxidative stress [23]. We also identified a number of genes that are essential for repairing damage to DNA such as that caused by reactive oxygen, including uvrA, uvrB, recB, ssb, mutM, and ruvC [24] (Figure 3B). This result is consistent with a screen for attenuated F. novicida U112 transposon mutants using an inhalation method of inoculation, which identified the DNA repair proteins recO and recA [21].
The fly's melanization immune response produces reactive oxygen as an effector and thus we hypothesized that these bacterial genes were involved in helping F. novicida resist melanization [25], [26]. To test this idea, we first performed in vitro disc diffusion assays to determine the sensitivity of each mutant to hydrogen peroxide (H2O2) and paraquat [27]. The oxyR mutants were extremely sensitive to both H2O2 and paraquat (Figure 3C (H2O2) and data not shown (paraquat)). In addition, all of the DNA damage repair mutants showed a significant degree of sensitivity to both reactive oxygen-producing agents (Figure 3D). Taken together, these data suggest that we identified a class of F. novicida genes that are essential for wild-type growth and survival within the fly, genes which help the bacteria to resist oxidative stress. Interestingly, 4 DNA repair genes, uvrA, recB, recO, and uvB, were identified in one screen of Francisella mutants in mice, suggesting that reactive oxygen species are a threat to the bacteria in mammalian infections as well.
To determine whether melanization is a critical factor limiting the growth of F. novicida, we performed competition assays using the oxyR mutant in CG3066 mutant flies. These flies do not induce melanization upon infection [17]. We found that the growth defect of oxyR with respect to wild type bacteria was rescued in non-melanizing CG3066 flies (Figure 3E). This supports the idea that melanization is the reactive oxygen producing immune response for limiting the growth of F. novicida in the fly. We therefore took our collection of negatively selected mutants and tested them for sensitivity to reactive oxygen. We found 25 mutants with increased sensitivity to oxidative stress and 2 with decreased sensitivity (Figure 2). Thus we were able to assign functions to these genes based on their behavior in a fly pathogenesis screen. This group of mutants makes up 45 percent of the mutants with attenuated growth phenotypes in the fly, demonstrating that oxidative stress mediated immunity is an important aspect of the fly's defenses against this pathogen and that an important class of F. novicida virulence factors in fly infections are genes that help the bacteria to counteract the effects of reactive oxygen.
Identification of F. novicida genes involved in bacterial resistance to imd-regulated innate immune responses
Having demonstrated that the TraSH method was useful for identifying genes required for growth in the fly and that many of these mutants were involved in counteracting the oxidative stress response of the fly, we decided to expand our study to examine an additional immune pathway and look for similar interactions. We chose to focus on one of the most intensely studied aspects of the fly innate immune response, NF-κB signaling pathways. Drosophila has two well-characterized NF-κB pathways (Toll and imd) that are responsible for sensing the presence of microbes and inducing an immune response [7]. Previous work by others demonstrated that the imd pathway is an important component of the Drosophila innate immune response to the LVS strain of F. tularensis, while the Toll pathway is not [9]. To confirm this for F. novicida U112 strain, we infected flies with null mutations in Toll and imd pathway genes. Two separate alleles of imd, imd1 and the null allele imd10191, as well as mutants lacking the NF-κB homologue Relish showed significantly increased sensitivity to infections with the U112 strain; in contrast, mutants in the Toll pathway components Dif1 and dMyD88C03881 showed no significant difference compared to wild type (Figure 4A (imd alleles only, relish data not shown) and Figure S1). Therefore, we focused on F. novicida genes required to resist the fly's imd mediated response.

To identify such genes, we repeated our TraSH analysis, this time infecting imd mutant flies and compared the results to those found in wild-type flies. We identified 36 genes that appeared to be negatively selected in wild type flies and at least partially rescued in imd mutant flies (Table S2) Subsequent confirmation of these results by competition assay using transposon insertion mutants revealed a subset of 7 mutants that showed reproducible large rescue phenotypes in imd flies (Figure 4C). These genes were the orphan response regulator pmrA, the gene FTN_0889 which is a helix-turn-helix protein and putative transcriptional regulator, glpD which is an anaerobic glycerol-3-phosphate dehydrogenase, the nicotinate-nucleotide pyrophosphorylase nadC, a uridine phosphorylase udp, FTN_0649, a FAD-dependent 4Fe-4S ferrodoxin, and FTN_0869, a hypothetical protein that encodes a putative transglutaminase that is regulated by the virulence factor mglA [28].
Since the imd pathway has been well-characterized as being responsible for inducing antimicrobial peptide (AMP) mRNA levels in response to F. novicida and other bacterial pathogens, the simplest explanation for the rescue of these bacterial mutants in flies lacking an intact imd pathway is that they have increased sensitivity to AMPs. This idea is further supported by identification of pmrA in our rescue screen, as pmrA has previously been shown to be sensitive to the antimicrobial peptide polymyxin B in vitro [29]. Therefore, we wished to determine whether the other F. novicida genes identified in our rescue screen are also sensitive to AMPs. There are 7 families of AMPs in Drosophila, and more than 2 dozen individual AMPs can be expressed during an infection, producing a complex bacteriocidal cocktail. Among the characterized AMPS, four families have been implicated in killing Gram-negative microbes, attacin, cecropin, diptericin, and drosocin. The first three families contain cation rich peptides while drosocin is described as proline rich [7]. It is currently impossible to recreate in vitro, the array of AMPs brought to bear on an infecting microbe in vivo. We therefore tried testing individual AMPs for their effects on F. novicida mutants. Unfortunately, few of these Drosophila AMPs are available commercially. We tested a commercial preparation of cecropin and did not detect activity against F.novicida on plates (data not shown). We turned to the cationic antimicrobial peptide polymyxin B, which has been used to model AMP sensitivity in F. tularensis in multiple studies [29], [30].
Of the seven genes we confirmed to be rescued in imd mutant flies, we found that five of these genes, pmrA, FTN_0889, glpD, udp, and FTN_0649 were indeed more sensitive to polymyxin B in vitro. Suprisingly, mutants in the genes FTN_0869 and nadC did not show any phenotypes in these assays, suggesting that the imd rescue phenotype of these mutants may not be due AMP sensitivity, or at least not to cationic AMP sensitivity (Figure 4D). To determine how common this phenotype was, we expanded our analysis to include the entire set of confirmed attenuated mutants described in Figure 2. We found that twelve of the fourteen F. novicida mutants that were rescued in imd mutant flies on arrays showed altered sensitivity to polymyxin B, whereas this was the case with just five of the thirty eight mutants not rescued in an imd mutant (Figure 2). These five mutants were likely exceptions as they also had defects in reactive oxygen sensitivity and in the absence of an imd mediated response would still be sensitive to a melanization response. In our entire set of attenuated mutants, only nadC and FTN_0869 mutants demonstrated the unique phenotype of rescue in an imd mutant fly without showing any increased sensitivity to AMPs, so we chose to focus on one of these genes, the putative transglutaminase FTN_0869 for further analysis.
FTN_0869 deletion mutants are attenuated in wild-type flies due to clearance of extracellular bacteria by an imd-dependent immune response
The mutation in the gene FTN_0869 was intriguing as it clearly grows better in imd mutants as compared to wild type flies yet the mutant does not demonstrate altered sensitivity to the antimicrobial peptide we tested. The fly produces dozens of AMPs at once and not all of them work by the same mechanism, therefore it is impossible and illogical to eliminate the possibility that a single untested AMP or combination of imd induced AMPs might be responsible for killing F. novicida. Regardless, the resistance of FTN_0869 mutants to an AMP raises the question that the imd pathway might be generating an immune response that was not AMP mediated. In addition, the fact that this gene is regulated by the virulence factor mglA which regulates the F. tularensis pathogenicity island and many other important virulence factors suggested that it could be particularly important to F. tularensis pathogenesis. To determine the extent of the attenuation of FTN_0869 mutants, we examined the growth and survival of these bacteria in individual infections.
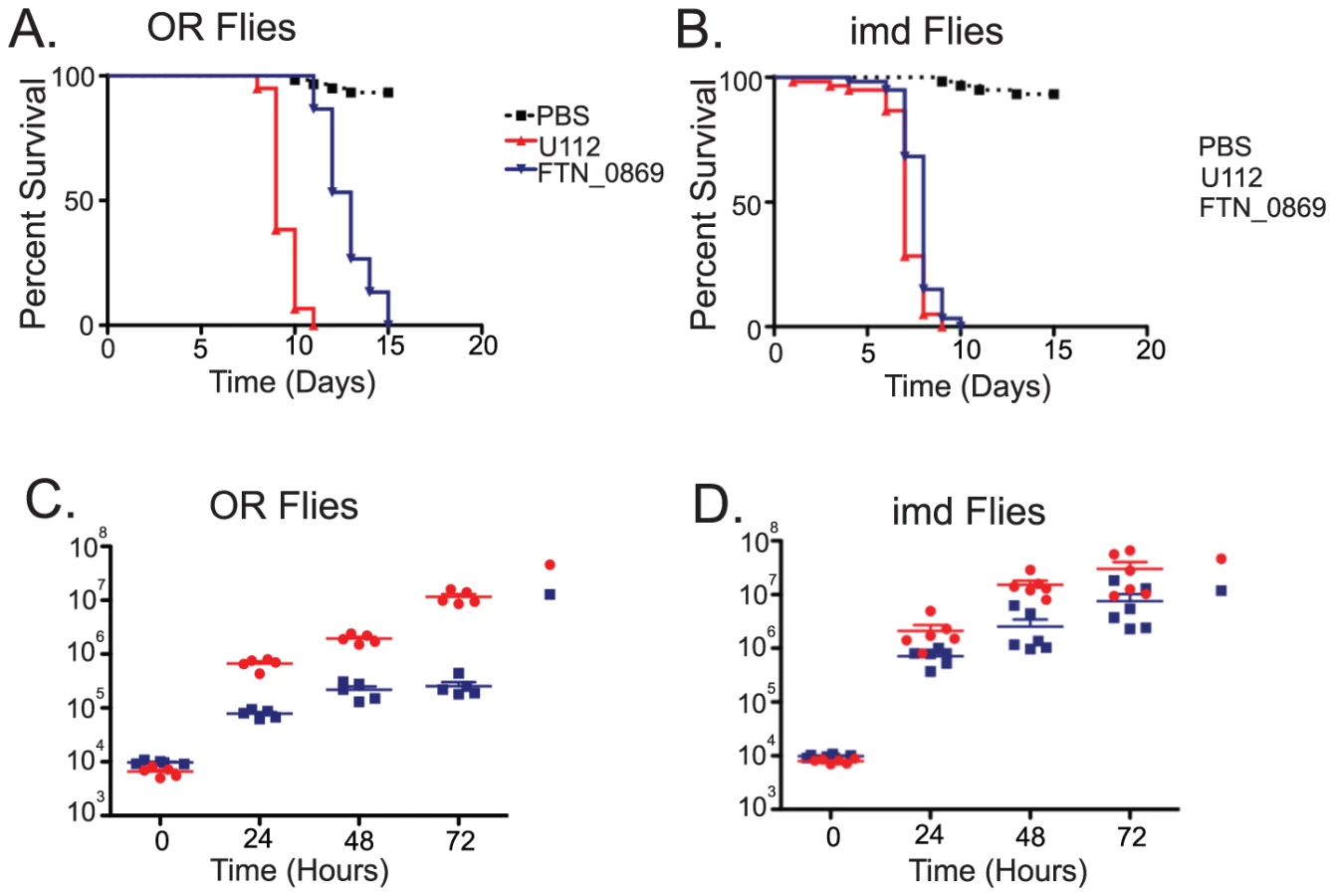
We observed that with a starting dose of 5×103 bacteria the FTN_0869 mutant took significantly longer to kill wild-type flies than did wild-type F. novicida (Figure 5A). This phenotype was completely rescued in imd mutant flies, with both FTN_0869 mutants and wild-type bacteria killing the fly with a mean time to death of 7 days, consistent with the sensitivity phenotype observed for imd flies (Figure 5B and Figure S1). In wild-type Drosophila, the FTN_0869 mutant did not develop the high bacterial loads found in wild type flies; wild type F. novicida can reach titers of 5×107 CFU per fly within 4 days while the FTN_0869 mutant did not grow higher than 5×105 CFU/fly (Figure 5C). Again, this phenotype was abrogated in imd mutant flies, in which both wild type bacteria and FTN_0869 mutants were able to grow to similar high titers. (Figure 5D) This suggested that an imd-regulated immune response was preventing the FTN_0869 mutants from growing as well as wild-type U112 bacteria in the fly. We infer that the decreased bacterial population was responsible for the decreased virulence observed in terms of fly survival.
The attenuated phenotype of mglA mutants in mouse cells is due to the inability of these mutants to survive and replicate intracellularly [31]. Since FTN_0869 is regulated by mglA, we sought to determine whether the same was true for this mutant. We performed gentamycin chase assays on wild-type U112, mglA mutants, and FTN_0869 mutants. As expected, the mglA mutants showed no bacterial growth within the fly but rather were partially cleared very quickly following injection into the fly, and were completely unable to establish an intracellular population (Figure 6A). This suggests that the small intracellular population may be important, if not essential, for the establishment of a successful infection. In contrast, the FTN_0869 mutants had a robust albeit slightly reduced intracellular population as compared to wild type bacteria, but demonstrated a unique phenotype with little to no extracellular bacteria present in wild-type flies (Figure 6B). By testing sensitivity to gentamycin in vitro, we were able to show that this was due to lack of extracellular bacteria rather than an increased sensitivity of the FTN_0869 mutant to gentamycin (Figure S3). Again, loss of the imd pathway in the host animal eliminated this effect; the extracellular population of FTN_0869 mutant bacteria grew to similar levels as wild-type bacteria in imd mutant flies (Figure 6C).

This result suggested that the extracellular population of bacteria was unable to persist in the extracellular space of infected flies due to an immune mechanism that is controlled by the imd signaling pathway. Therefore, we were interested in investigating this mutant further to determine what effector arm of the innate immune system was responsible for the clearance of extracellular bacteria. We hypothesized that this clearance could be due to either an increased activation of the imd pathway by the FTN_0869 mutants, AMP activity that we were unable to test in vitro, or a novel component of the imd-regulated immune response. To determine if the imd pathway is induced more intensely by the FTN_0869 mutant, to rule out the possibility that this gene is able to downregulate the imd immune response, we measured the induction of imd-regulated AMPS as a readout for imd pathway activation. We used quantitative real-time RT-PCR to monitor the levels of Diptericin, Drosocin, Drosomycin, Attacin, Cecropin and Metchnikowin at 1,2,5 and 24 hours post-infection. We found that only Metchnikowin, Cecropin, and Diptericin were strongly induced in response to F. novicida infections and that the transcript levels of each of these highly-induced AMPs peaked at 24 hours post-infection (Figure 7A and data not shown). All of these AMPs were induced to similar levels during infections with either the wild-type or FTN-0869 mutant bacteria, with no statistically significant difference between induction by wild-type or mutant bacteria at any timepoint. This confirms that the imd pathway is indeed activated by F. novicida and that the gene FTN_0869 does not have an effect on the induction of the imd pathway.

Clearance of extracellular FTN_0869 mutants is not dependent on antimicrobial peptides or melanization
We wished to probe the role of AMPs in clearing F. novicida further. There are more than 30 antimicrobial peptides in the fly and purified Drosophila AMPs are not readily available and the AMPs are always expressed together during an immune response; as described above, this makes it difficult to directly test the role of AMP activity on F. novicida growth in the fly. We therefore tried an indirect approach to test their importance. Recent work in the beetle Tenebrio molitor demonstrated that the majority of bacteria injected into the insect is cleared in less than an hour post-infection, much faster than antimicrobial peptides can be upregulated, transcribed, and synthesized [32]. Using this larger insect model, Haine et al. were able to conclusively demonstrate that insect antimicrobial peptide activity is induced slowly, and thus is not responsible for the bulk of the bacterial clearance. The analysis of antimicrobial peptide induction in the fly relies on the analysis of mRNA transcript levels, which are less accurate kinetically than a direct measurement of antimicrobial activity but nevertheless suggest that a slow induction with transcript levels only rising hours after infection and peaking at 6–24 hours post-infection for various AMPs [33]. To determine if the kinetics of extracellular bacterial clearance coincide with AMP induction, we performed gentamycin chase assays at early timepoints post infection. As early as 1 hour post-infection much of the FTN_0869 mutant population had already been cleared from the fly; in contrast the extracellular population of wild-type bacteria did not substantially decrease (Figure 7B). By two hours post-infection, the timepoint at which both wild-type and mutant bacteria had begun to enter cells, the wild-type bacteria now had both intracellular and extracellular populations, while in FTN_0869 mutant infections only the intracellular bacteria had survived clearance. By five hours post infection the extracellular population of U112 wild-type bacteria had begun to increase while the titer of FTN_0869 mutants did not and only the intracellular population remained. This supported the notion that imd-induced AMPs were not responsible for the clearance of extracellular FTN_0869 mutant bacteria, as the bulk of this clearance occurred within an hour post-infection before AMP activity would be upregulated.
We next sought to determine if one of the other effector arms of the fly innate immune system could be causing this phenotype. We first examined the effects of reactive oxygen species on the FTN_0869 mutants. Unlike many of the genes we isolated from our TraSH screen, the FTN_0869 mutants did not show increased sensitivity to reactive oxygen species produced by H2O2 or paraquat in vitro as measured by disk diffusion assay (Figure 2). We next examined the effects of melanization in vivo by infecting CG3066 mutant flies with wild type and FTN_0869 mutants. Unlike the oxyR mutants, the FTN_0869 mutants were just as attenuated in CG3066 mutants as they are in the wild-type control (Figure 7C) suggesting that these mutants do not have a defect in resisting reactive oxygen stress and that melanization is not responsible for the FTN_0869 imd rescue phenotype. We concluded that the imd rescue phenotype of FTN_0869 mutants not likely due to cationic antimicrobial peptides or melanization and rather suggested a third category of F. novicida interactions with the fly immune system as shown in Figure 2.
Discussion
Our goal was to dissect the host-pathogen interactions between Francisella and D. melanogaster. To identify components of this complex system we used a combination of three genetic techniques that enabled us to determine the contributions of both host and microbe to the virulence of the infection. First, we identified bacterial virulence factors necessary to infect the fly using a library of F. novicida mutants. Second, we used fly immunity mutants to confirm which host immune pathways were essential for fighting F. novicida infections. Finally, we combined these two techniques to identify subsets of bacterial virulence factors that allow the bacteria to counter-respond to specific immune attacks and evade immune clearance. This paper identifies genes from both the pathogen and the host that are components of each of these aspects of the host-pathogen relationship.
To identify bacterial virulence factors, we performed an in vivo screen that identified 149 bacteria genes that are important for growth and survival within the fly. 41 of the 149 genes had previously been identified in a similar screen performed with the same bacterial library in the mouse indicating that many bacterial virulence factors are conserved between host species [20]–[22]. Genes that overlap between the Drosophila and mouse screens include known virulence factors such as mglA, iglC, and iglD, a number of various transporters, and some of the DNA repair genes we identified as helping F. novicida to survive oxidative stress. The remaining genes are unique to our screen performed in the fly model. These genes could either represent F. novicida genes that play a role specific to arthropod vectors, demonstrate a stronger phenotype in insects than in mammals, or were not identified in previous screens for experimental reasons.
We note that of the 26 F.novicida mutants identified as being sensitive to reactive oxygen, 7 (27%) had been previously identified as being important for virulence in vertebrates. In contrast, of the 16 mutants we found to be polymixin sensitive, only 1 (7%) was identified previously as being important for virulence in vertebrates. The numbers in this study are small enough that differences in representation could be due to chance and therefore future work with more pathogens will be required to confirm the trends seen here; that said, analysis of interactions with the fly's reactive oxygen based immune response seems to be useful predictor of genes that will be of interest to those studying vertebrates. In contrast, analysis of the AMP and imd sensitive mutants is not as robust a tool for identifying mutations that will are relevant in vertebrates.
Secondary screens of these mutants revealed important patterns that shed light onto what particular stresses F. novicida encounters within the fly. 25 of the 56 mutants that we confirmed to have reduced competitive indexes compared to wild-type F. novicida were also hyper sensitive to oxidative stress in vitro. This indicates that preventing or repairing damage caused by reactive oxygen species is an important survival strategy for F. novicida in insect infections. Of particular interest among the genes that were sensitive to oxidative stress was the gene oxyR, which has homology to an E. coli transcriptional regulator that senses and responds to the presence of hydrogen peroxide by inducing the transcription of catalases and other genes that can counteract oxidative stress 23. In addition, our screen identified multiple genes in DNA damage repair pathways that are also sensitive to oxidative stress [24], [25]. We expect that these genes are required to repair damage caused by reactive oxygen species to DNA as has been suggested by Kraemer et al [21].
Of the three effector arms that have been characterized in the immune response occurring within the fly's body cavity, the major producer of oxidative stress is the melanization response [15], [17], [26]. Therefore, we speculated that the large number of negatively selected bacterial mutants with oxidative stress sensitivity phenotypes suggested that the melanization response plays a large role in the fly's immune response to F. novicida. To test this hypothesis, we performed competition assays with the oxyR mutants in fly mutants that lack a melanization response. As expected, the attenuation of these mutants was rescued in flies that do not melanize and therefore would be expected to not produce toxic oxygen species. This demonstrated that melanization is an essential component of the fly immune response against F. novicida and demonstrated that we could use our characterizations of bacterial genes to learn about the fly immune system and understand the host-pathogen relationship. We note that reactive oxygen is a well-established immune effector in the Drosophila gut. Perhaps most microbes encountering the fly will face this immune barrier before encountering internal immune defenses. Thus protection against reactive oxygen is doubly important for fly pathogens [34].
Because microbes must withstand the host immune system to mount a successful infection, we were able to exploit the inherent ability of bacteria to function as metaphorical immunologists to identify which aspects of fly immunity were important to F. novicida infections. We next sought to determine if this system could be used in the reverse direction by manipulating the fly immune system to identify which bacterial virulence factors were responsible for interacting with one specific aspect of innate immunity. We did this by performing a second round of our in vivo screen for bacterial mutants in an immunocompromised fly. We focused on the imd-regulated humoral immune response, which had previously been identified as important for fighting F. tularensis infections [9]. We confirmed that the imd pathway, but not the Toll pathway, was essential in combating F. novicida infections, and performed our TraSH assay in imd mutant flies. We identified a subset of bacterial virulence factors that were important for infections of wild-type flies but not imd flies; this imd-regulated immune response has been primarily characterized for its role in the induction of antimicrobial peptides and therefore we tested these mutants for their sensitivity to a cationic, membrane active antimicrobial peptide, polymyxin B [29], [30]. As expected, twelve of the fourteen mutants were sensitive to polymyxin killing in vitro, providing another example of how resistance to host immune responses is an important component of bacterial virulence.
We identified 2 bacterial mutants that were not sensitive to polymyxin in vitro despite being rescued in imd mutants flies. This phenotype was unexpected as the majority of the literature suggests imd signaling drives antimicrobial peptide production and this is its most important job. We propose three explanations for this phenotype. First, the rescue phenotype could be due to specific sensitivity to additional antimicrobial peptides that were not tested in vitro; the bacteria show no sensitivity to polymyxin but could be sensitive to one of the 30 or more AMPs synthesized by flies. Second, the rescue phenotype could be due to the bacterial gene being an inhibitor of the imd pathway; in this case the bacteria would have wild type sensitivity to AMPs but would encounter increased concentrations of them in the fly because the bacteria could not inhibit AMP production. Finally, the rescue phenotype of these bacterial mutants could be due to an aspect of imd-regulated immunity that has not been previously described.
To differentiate between these possibilities, we chose one imd-rescue mutant, the putative transglutaminase FTN_0869 to characterize further in terms of its interactions with the fly immune system. We chose this gene because it had a strongly attenuated phenotype in wild-type flies that was significantly rescued in imd mutants and because it has previously been shown to be regulated by the virulence factor mglA, which is essential for F. novicida intracellular growth 28. More recently, the homologue of this gene in the extremely virulent Type A F. tularensis ssp. tularensis strain Shu4 was identified in a transcriptional analysis of genes that are upregulated inside mouse bone marrow-derived macrophages (BMMs) [35]. Interestingly, FTN_0869 deletion mutants in the less virulent U112 strain are unable to replicate in BMMs, but mutants of the homologue of this gene, FTT0989 in the SCHU4 strain did not demonstrate any intracellular replication defect [28], [35]. Further characterization of the FTN_0869 mutants showed that these mutants are attenuated for both lethality to the fly and bacterial growth in an imd-dependent manner. However, unlike its transcriptional regulator mglA, the FTN_0869 mutant is capable of intracellular growth within flies, but is incapable of surviving in the extracellular space. This phenotype is consistent with what is observed in mouse bone marrow-derived macrophages with the virulent Shu4 strain, but not with the phenotype of FTN_0869 deletion mutants in the U112 strain. The reason for this difference is unclear, but it is interesting to note that the ability of the putative transglutaminase deletion mutants to grow intracellularly correlates with its virulence in mammalian and insect hosts.
We found that the phenotype of FTN_0869 deletion mutants in flies is imd-dependent, and used this phenotype to investigate the role of the imd pathway in clearing the extracellular bacteria. With this mutant, we were able to show that the imd rescue phenotype of this particular mutant was not due to modulation of the imd pathway because AMP genes downstream of imd are induced to similar amounts in infections with wild-type and FTN_0869 bacteria. By examining the kinetics of the clearance of extracellular bacteria, we were able to limit the possibility that other imd-induced antimicrobial peptides that we did not test in vitro were causing the attenuation of the FTN_0869 mutant. Using non-melanizing mutants, we were able to rule out melanization as the cause of this phenotype, leaving us with the possibility that imd could either be regulating the cellular immune response or an uncharacterized effector arm of fly immunity. Thus the FTN_0869 phenotype suggested a third category of host-pathogen interactions between F. novicida and the Drosophila innate immune system. Future work with this mutant and other imd-rescue mutants identified in our screen could provide further insight into the biology of the imd-regulated fly immune response.
In summary, reciprocal studies of a pathogen, F. novicida and a host, D. melanogaster, allowed us to identify genes in the pathogen required to counteract, evade, or resist host immune responses and allow bacterial growth and survival. These studies identified two branches of host immunity that are important for fighting F. novicida infections, melanization and imd-regulated immune responses and helped us to understand how the bacteria resists these responses. By identifying the mechanism of one or two bacterial mutants based on their sequence or interaction with fly mutants we developed assays to identify the mechanism of mutants with unknown function. Our work with one of these mutants, FTN_0869, taught us that there is likely more to learn about the fly immune system as there are classes of F. novicida mutants that cannot immediately be explained by their interactions with the melanization response or AMPs. Our screen allowed us to pose directed questions and focus our investigations on particular aspects of the host immune system and the microbial strategies to evade this immune response, helping us to identify and characterize components of the host-pathogen relationship.
Materials and Methods
Drosophila strains
All experiments were performed in wild-type Oregon Red (OR) flies unless otherwise noted. The imd mutant fly line imd10191 is a null allele with a 26-nucleotide deletion at amino acid 179 that results in a frameshift mutation and has been backcrossed onto an OR background. The Toll pathway alleles tested in this study are Dif1 which is a complete loss of function mutant and MyD88C03881 The CG3066KG02818 Sp7 mutant flies are PiggyBack insertion mutants on a w1118 background (Bloomington stock number 13494), and w1118 flies are used as the wild-type control for these experiments. All experiments were performed on 5–7 day old age-matched male flies that were maintained on dextrose medium at 25°C and 65% humidity in a 12∶12h light dark cycle.
Bacterial cultures
Francisella novicida strain U112 was used for all experiments described in this paper. Bacterial stocks were grown in Tryptic Soy Broth (TSB) supplemented with 0.2% L-cysteine and cultured overnight under aerobic conditions at 37°C. Cultures were grown to an OD600 of 1.5–2 and diluted in PBS to OD600 0.005–0.01 for fly infections.
Fly infections
Flies were anaesthetized with CO2 and injected with 50nL of bacteria using a glass needle and a Picospritzer III injector system (Parker Hannifin). Each fly was injected in the ventrolateral surface of the fly abdomen and placed into fresh vials with no more than 20 flies per vial to prevent crowding. Following infection, the flies were incubated at either 25°C or 29°C as noted. Each survival curve was performed using 3 replicates of 20 flies each for a total of 60 flies per condition and each experiment was performed a minimum of three times. The number of dead flies was monitored daily and Kaplan-Meier survival curves were generated using GraphPad Prism software, and statistical analysis was performed using log-rank analysis.
Determination of bacterial CFUs and gentamycin-chase assays
Individual infected flies were homogenized in 100µL of PBS, serially diluted, and plated onto Mueller-Hinton (MH) agar plates supplemented with 0.025% ferric pyrophosphate (Sigma), 0.1% glucose, 0.025% calf serum (GIBCO), and 0.02% Iso-VitaleX (Benton Dickinson). Plates were incubated overnight and colonies were counted to determine the number of bacterial colony forming units (CFUs) per fly. Statistical significance was determined using unpaired two-tailed t-tests. Gentamycin chase experiments were performed as described about except that 50nL of 1mg/mL of gentamycin was injected into each fly 3 hours prior to plating [36].
TraSH experiments
Three sets of 30 flies were injected with 50nL of the trash library. Each fly received approximately 2*105 CFUs of bacteria, representing approximately 2-fold coverage of the library. The infection was allowed to proceed for two days at 29°C, at which point each fly was homogenized and plated onto MH agar. Plates were incubated at 37°C overnight, and the bacteria were collected and pooled and DNA was collected by phenol-chloroform extraction. Each pool was divided in half and digested with either BfaI or RsaI (NEB) and re-pooled to be used as a template for in vitro transcription with a MegaScript T7 Kit (Ambion). The RNA was then purified and used for reverse transcription using a SuperScript III First Strand Synthesis Kit (Invitrogen) and random hexamer primers. The resulting cDNA was labeled with amino-allyl dUTP using Klenow (exo-) enzyme (NEB). The input pool was then labelled with Cy5 and the day 2 pools with Cy3 and hybridized to Francisella microarrays as has been previously described [28]. Data was normalized using the Stanford Microarray Database according to the median log2 Cy5/Cy3 and filtered using a Cy3 net median intensity of 150 and a regression correlation of >0.6. The dataset was then analyzed using SAM software using a blocked 2-class analysis to identify differences between the input and wild-type or input and imd mutant samples with a false discovery rate of 5% [37].
Construction of bacterial mutants
Genes that were selected for further analysis were knocked out of F. novicida individually to create deletion mutants. Briefly, 500bp of sequence 5′ and 3′ to the gene of interest was amplified from genomic F. novicida DNA using Phusion DNA Poylmerase (NEB), and fused onto either side of a kanamycin cassette using a sewing PCR reaction. 38 The resulting PCR products were then transformed into chemically competent F. novicida U112 as described [28] and the mutants were confirmed by PCR.
Competition assays
To confirm the bacterial growth attenuation phenotypes, 50nL of a 1∶1 ratio of mutant and wild-type bacteria at an OD600 of 0.01 was injected into flies. The infection was allowed to proceed for 2 days at 29°C, following which the flies were homogenized and plated onto MH agar plates with and without 30 mg/mL of kanamycin. Since only the mutant bacteria is capable of growing in kanamycin media, we were able to determine the number of wild-type and mutant bacterial CFUs for each fly by subtracting the number of mutant bacterial CFUs from the total CFUs per fly. A competitive index (CI) was determined using the formula CI = (mutant CFU day 2/wild-type CFU day 2)/(mutant CFU input/wild-type CFU input).
Disk diffusion assays
To determine the sensitivity of various F. novicida mutants to oxidative stress and antimicrobial peptides, disk diffusion assays were performed using protocols adapted from Mohapatra et al. and Bakshi et al. [27], [29]. Briefly, 50µL of overnight cultures of bacteria were plated onto MH agar plates to create a lawn of bacteria. Plates were allowed to dry for 10 minutes, and then 6mm Whatman filter paper disks (Fisher Scientific) were placed onto each plate and inoculated with 10µL of 100mM freshly diluted hydrogen peroxide (Sigma) or 10µL of a 10 mg/mL stock of polymyxin B. Plates were incubated overnight and the diameter of the zone of inhibition was measured for each sample. Three zones were measured for each mutant and each experiment was repeated three times.
qRT-PCR of antimicrobial peptides
The fold increase of antimicrobial peptide expression following infection by wild-type and FTN_0869 mutant F. novicida was determined by isolating RNA from infected flies 6 and 24 hours post-infection by trizol extraction and performing qRT-PCR analysis using an iScript One-Step RT-PCR kit with SYBR Green (Bio-Rad) and a Bio-Rad icycler. The following primer sets were used: cecropin 5′ 5″-tcttcgttttcgtcgctctc-3′, cecropin 3′ 5′-cttgttgagcgattcccagt-3′, drosomycin 5′ 5′-gacttgttcgccctcttcg-3′, drosomycin 3′ 5′-cttgcacacacgacgacag-3′, diptericin 5′ 5′-accgcagtacccactcaatc-3′, diptericin 3′ 5′-cccaagtgctgtccatatcc-3′, attacin 5′ 5′-caatggcagacacaatctgg-3′, attacin 3′ 5′-attcctgggaagttgctgtg-3, drosocin 5′ 5′-ttcaccatcgttttcctgct-3′, drosocin 3′ 5′-agcttgagccaggtgatcct-3′, metchinkowin 5′ 5′-tcttggagcgatttttctgg3′, metchnikowin 3′ 5′aataaattggacccggtcttg-3′, ribosomal protein 15a 5′-tggaccacgaggaggctagg, 3′-gttggttgcatcctcggtga.
Supporting Information
Zdroje
1. FinlayBB
FalkowS
1989 Common themes in microbial pathogenicity. Microbiol Rev 53 210 230
2. SchneiderDS
AyresJS
2008 Two ways to survive infection: what resistance and tolerance can teach us about treating infectious diseases. Nat Rev Immunol 8 889 895
3. PerssonJ
VanceRE
2007 Genetics-squared: combining host and pathogen genetics in the analysis of innate immunity and bacterial virulence. Immunogenetics 59 761 778
4. LemaitreB
NicolasE
MichautL
ReichhartJM
HoffmannJA
1996 The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86 973 983
5. RosettoM
EngstromY
BaldariCT
TelfordJL
HultmarkD
1995 Signals from the IL-1 receptor homolog, Toll, can activate an immune response in a Drosophila hemocyte cell line. Biochem Biophys Res Commun 209 111 116
6. Shirasu-HizaMM
SchneiderDS
2007 Confronting physiology: how do infected flies die? Cell Microbiol 9 2775 2783
7. UvellH
EngstromY
2007 A multilayered defense against infection: combinatorial control of insect immune genes. Trends Genet 23 342 349
8. AggarwalK
SilvermanN
2008 Positive and negative regulation of the Drosophila immune response. BMB Rep 41 267 277
9. VonkavaaraM
TelepnevMV
RydenP
SjostedtA
StovenS
2008 Drosophila melanogaster as a model for elucidating the pathogenicity of Francisella tularensis. Cell Microbiol 10 1327 1338
10. NigrovicLE
WingerterSL
2008 Tularemia. Infect Dis Clin North Am 22 489 504, ix
11. PetersenJM
MeadPS
SchrieferME
2009 Francisella tularensis: an arthropod-borne pathogen. Vet Res 40 7
12. 2005 Tularemia transmitted by insect bites–Wyoming, 2001–2003. MMWR Morb Mortal Wkly Rep 54 170 173
13. HankeCA
2009 Ulceroglandular tularemia in a toddler in Germany after a mosquito bite. Eur J Pediatr 168 937 940
14. KlockLE
OlsenPF
FukushimaT
1973 Tularemia epidemic associated with the deerfly. JAMA 226 149 152
15. AyresJS
SchneiderDS
2008 A signaling protease required for melanization in Drosophila affects resistance and tolerance of infections. PLoS Biol 6 2764 2773
16. LeclercV
PelteN
El ChamyL
MartinelliC
LIgoxygakisP
2006 Prophenoloxidase activation is not required for survival to microbial infections in Drosophila. EMBO Rep 7 231 235
17. TangH
KambrisZ
LemaitreB
HashimotoC
2006 Two proteases defining a melanization cascade in the immune system of Drosophila. J Biol Chem 281 28097 28104
18. BarkerJR
KloseKE
2007 Molecular and genetic basis of pathogenesis in Francisella tularensis. Ann N Y Acad Sci 1105 138 159
19. ReadA
VoglSJ
HuefferK
GallagherLA
HappGM
2008 Francisella genes required for replication in mosquito cells. J Med Entomol 45 1108 1116
20. WeissDS
BrotckeA
HenryT
MargolisJJ
ChanK
2007 In vivo negative selection screen identifies genes required for Francisella virulence. Proc Natl Acad Sci U S A 104 6037 6042
21. KraemerPS
MitchellA
PelletierMR
GallagherLA
WasnickM
2009 Genome-wide screen in Francisella novicida for genes required for pulmonary and systemic infection in mice. Infect Immun 77 232 244
22. SuJ
YangJ
ZhaoD
KawulaTH
BanasJA
ZhangJR
2007 Genome-Wide Identification of Francisella tularensis Virulence Determinants. Infect Immun 75 3089 3101
23. MongkolsukS
HelmannJD
2002 Regulation of inducible peroxide stress responses. Mol Microbiol 45 9 15
24. NowosielskaA
2007 Bacterial DNA repair genes and their eukaryotic homologues: 5. The role of recombination in DNA repair and genome stability. Acta Biochim Pol 54 483 494
25. ImlayJA
2002 How oxygen damages microbes: oxygen tolerance and obligate anaerobiosis. Adv Microb Physiol 46 111 153
26. NappiA
PoirieM
CartonY
2009 The role of melanization and cytotoxic by-products in the cellular immune responses of Drosophila against parasitic wasps. Adv Parasitol 70 99 121
27. BakshiCS
MalikM
ReganK
MelendezJA
MetzgerDW
2006 Superoxide dismutase B gene (sodB)-deficient mutants of Francisella tularensis demonstrate hypersensitivity to oxidative stress and attenuated virulence. J Bacteriol 188 6443 6448
28. BrotckeA
WeissDS
KimCC
ChainP
MalfattiS
2006 Identification of MglA-regulated genes reveals novel virulence factors in Francisella tularensis. Infect Immun 74 6642 6655
29. MohapatraNP
SoniS
BellBL
WarrenR
ErnstRK
2007 Identification of an orphan response regulator required for the virulence of Francisella spp. and transcription of pathogenicity island genes. Infect Immun 75 3305 3314
30. WangX
RibeiroAA
GuanZ
AbrahamSN
RaetzCR
2007 Attenuated virulence of a Francisella mutant lacking the lipid A 4′-phosphatase. Proc Natl Acad Sci U S A 104 4136 4141
31. BaronGS
NanoFE
1998 MglA and MglB are required for the intramacrophage growth of Francisella novicida. Mol Microbiol 29 247 259
32. HaineER
MoretY
Siva-JothyMT
RolffJ
2008 Antimicrobial defense and persistent infection in insects. Science 322 1257 1259
33. LemaitreB
ReichhartJM
HoffmannJA
1997 Drosophila host defense: differential induction of antimicrobial peptide genes after infection by various classes of microorganisms. Proc Natl Acad Sci U S A 94 14614 14619
34. RyuJH
HaEM
LeeWJ
2010 Innate immunity and gut-microbe mutualism in Drosophila. Dev Comp Immunol 34 369 376
35. WehrlyTD
ChongA
VirtanevaK
SturdevantDE
ChildR
2009 Intracellular biology and virulence determinants of Francisella tularensis revealed by transcriptional profiling inside macrophages. Cell Microbiol 11 1128 1150
36. BrandtSM
DionneMS
KhushRS
PhamLN
VigdalTJ
2004 Secreted Bacterial Effectors and Host-Produced Eiger/TNF Drive Death in aSalmonella-Infected Fruit Fly. PLoS Biol 2 e418
37. TusherVG
TibshiraniR
ChuG
2001 Significance analysis of microarrays applied to the ionizing radiation response. Proc Natl Acad Sci U S A 98 5116 5121
38. LiuJ
ZogajX
BarkerJR
KloseKE
2007 Construction of targeted insertion mutations in Francisella tularensis subsp. novicida. Biotechniques 43 487 90, 492
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2010 Číslo 8
- Farmakovigilanční studie perorálních antivirotik indikovaných v léčbě COVID-19
- Jak souvisí postcovidový syndrom s poškozením mozku?
- Měli bychom postcovidový syndrom léčit antidepresivy?
- 10 bodů k očkování proti COVID-19: stanovisko České společnosti alergologie a klinické imunologie ČLS JEP
Nejčtenější v tomto čísle
- The Transcription Factor Rbf1 Is the Master Regulator for -Mating Type Controlled Pathogenic Development in
- PKC Signaling Regulates Drug Resistance of the Fungal Pathogen via Circuitry Comprised of Mkc1, Calcineurin, and Hsp90
- Contribution of Coagulases towards Disease and Protective Immunity
- Early Severe Inflammatory Responses to Uropathogenic Predispose to Chronic and Recurrent Urinary Tract Infection