
Integrative Genomic Analysis Identifies Isoleucine and CodY as Regulators of Virulence
Intracellular bacterial pathogens are metabolically adapted to grow within mammalian cells. While these adaptations are fundamental to the ability to cause disease, we know little about the relationship between the pathogen's metabolism and virulence. Here we used an integrative Metabolic Analysis Tool that combines transcriptome data with genome-scale metabolic models to define the metabolic requirements of Listeria monocytogenes during infection. Twelve metabolic pathways were identified as differentially active during L. monocytogenes growth in macrophage cells. Intracellular replication requires de novo synthesis of histidine, arginine, purine, and branch chain amino acids (BCAAs), as well as catabolism of L-rhamnose and glycerol. The importance of each metabolic pathway during infection was confirmed by generation of gene knockout mutants in the respective pathways. Next, we investigated the association of these metabolic requirements in the regulation of L. monocytogenes virulence. Here we show that limiting BCAA concentrations, primarily isoleucine, results in robust induction of the master virulence activator gene, prfA, and the PrfA-regulated genes. This response was specific and required the nutrient responsive regulator CodY, which is known to bind isoleucine. Further analysis demonstrated that CodY is involved in prfA regulation, playing a role in prfA activation under limiting conditions of BCAAs. This study evidences an additional regulatory mechanism underlying L. monocytogenes virulence, placing CodY at the crossroads of metabolism and virulence.
Published in the journal:
. PLoS Genet 8(9): e32767. doi:10.1371/journal.pgen.1002887
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002887
Summary
Intracellular bacterial pathogens are metabolically adapted to grow within mammalian cells. While these adaptations are fundamental to the ability to cause disease, we know little about the relationship between the pathogen's metabolism and virulence. Here we used an integrative Metabolic Analysis Tool that combines transcriptome data with genome-scale metabolic models to define the metabolic requirements of Listeria monocytogenes during infection. Twelve metabolic pathways were identified as differentially active during L. monocytogenes growth in macrophage cells. Intracellular replication requires de novo synthesis of histidine, arginine, purine, and branch chain amino acids (BCAAs), as well as catabolism of L-rhamnose and glycerol. The importance of each metabolic pathway during infection was confirmed by generation of gene knockout mutants in the respective pathways. Next, we investigated the association of these metabolic requirements in the regulation of L. monocytogenes virulence. Here we show that limiting BCAA concentrations, primarily isoleucine, results in robust induction of the master virulence activator gene, prfA, and the PrfA-regulated genes. This response was specific and required the nutrient responsive regulator CodY, which is known to bind isoleucine. Further analysis demonstrated that CodY is involved in prfA regulation, playing a role in prfA activation under limiting conditions of BCAAs. This study evidences an additional regulatory mechanism underlying L. monocytogenes virulence, placing CodY at the crossroads of metabolism and virulence.
Introduction
Intracellular bacterial pathogens have developed sophisticated mechanisms to enter eukaryotic cells and replicate within them. These mechanisms involve bacterial proteins that overcome host defense strategies and barriers as well as nutritional limitations. Intracellular pathogens are generally categorized into two groups based on the compartment where they replicate, the vacuole/phagosome or the host cell cytosol. Each intracellular niche presents unique nutritional challenges demanding that bacteria exhibit specific metabolic adaptations to proliferate successfully. Vacuolar pathogens, such as Legionella pneumophilla and Mycobacterium tuberculosis, actively modify their compartment via secretion of effector proteins that enrich the vacuole with nutrients to support growth; nevertheless growth rate in the vacuole is much slower than in rich media. In contrast, intracellular cytosolic bacteria, such as Listeria monocytogenes, Sheigella felxenri and Burkholderia pseudomallei manage to exploit their niche such that growth rates resemble growth in rich media [1]. Little is known about the metabolic adaptations that enable intracellular cytosolic pathogens to grow rapidly or if such adaptations affect virulence. A better understanding of how these bacteria overcome nutritional limitations will give insight into cytosol nutrient composition and could facilitate development of drugs against intracellular pathogens.
L. monocytogenes is a Gram-positive facultative intracellular bacterial pathogen and the causative agent of listeriosis in humans, a disease with a variety of clinical manifestations including meningitis and abortion [2]. L. monocytogenes infects phagocytic and non-phagocytic cells, using surface expressed proteins called internalins, which bind and induce bacterial uptake by endocytosis [3]. Upon entry, L. monocytogenes escapes from the phagosome/vacuole into the host cytosol by producing the pore-forming hemolysin toxin, listeriolysin O (LLO, encoded by the hly gene), and two additional phospholipases [4]–[6]. Once in the host cytosol, L. monocytogenes multiplies rapidly and expresses the surface protein, ActA, which recruits the host actin polymerization machinery to propel the bacteria in the cytosol and facilitate spread from cell to cell [7], [8]. All known virulence factors involved in internalization, vacuolar escape and cell-to-cell spread are co-regulated by the major virulence activator, PrfA [9].
L. monocytogenes uses several carbon sources during intracellular growth, but primarily glycerol, di-hydroxyacetone and phosphorylated carbohydrates (such as glucose 1-phosphate), indicating the availability of these substrates in the cytosolic niche [10]–[12]. Glycerol uptake is mediated by a glycerol permease, whereas phosphorylated sugars are transported via the specialized hexose-phosphate transporter, Hpt. Both systems are induced intracellularly and are important for bacterial replication [11], [13]. It is well established that carbon metabolism during intracellular growth is linked directly to the virulence of L. monocytogenes. Carbohydrates transported via the phosphoenolpyruvate phosphotransferase system (PTS) (such as glucose and cellobiose) repress the activity of PrfA whereas non-PTS carbon sources (such as glycerol and glucose 1-phosphate that are available in the host cytosol) induce PrfA activity, resulting in elevated expression of virulence genes [reviewed in: [14] and [15]]. Based on these observations, it was proposed that non-PTS sugars might serve to signal L. monocytogenes of its intracellular location.
Various additional metabolic pathways were indicated as important for intracellular replication of L. monocytogenes, including several amino acid biosynthesis pathways such as the branch chain amino acid (BCAA) and arginine pathways [16], [17], the common aromatic compounds biosynthesis (shikimate) pathway, as well as the synthesis and uptake of thiamine (vitamin B1) [18], [19]. A special adaptation of L. monocytogenes to the cytosolic niche is its ability to obtain the co-factor lipoate from the host, as it cannot be synthesized by the bacteria. Listerial expression of a lipoate ligase, LplA1, enables the co-factor to be derived from host lipoyl-peptides [20]. As for nitrogen sources, it is thought that L. monocytogenes utilizes ammonium, arginine and ethanolamine [21], [22]. The latter is highly abundant in mammalian cells as it is the breakdown product of phosphatidylethanolamine. The ability to use ethanolamine as a nitrogen and/or carbon source is linked to the pathogenesis of several bacteria, such as Salmonella, Enterococcus and Clostridium [22]. Like Salmonella, listerial genomes encode the ethanolamine utilization pathway [23]. Taken together, these various adaptations highlight the complex metabolic requirements of intracellular growth.
Recent advances in genomic sequencing and development of constraint-based metabolic models enable now the reconstruction of genome scale metabolic networks of different organisms. This approach enables a better comprehension of the complete metabolic network of an organism under different growth conditions as well as the prediction of essential metabolic genes [24], [25]. These remarkable advances have prompted several studies of the metabolic networks of pathogens during infection, with the goal of characterizing potential drug targets [26]–[30]. Lately, as part of the SEED project an automated genome-scale metabolic model of L. monocytogenes metabolism has been developed [31] that comprises a stoichiometric matrix of reactions and metabolites representing the organism's entire metabolic network. In this study we used this metabolic model to analyze the global metabolic state of L. monocytogenes during infection.
To model the metabolic state of an organism under a given set of conditions, the availability of nutrients needs to be simulated. This information is mostly unknown for complex systems, but a specific metabolic state can be specified by integrating relevant transcriptome, proteome or metabolome (‘omics’) data into the metabolic model. A computational tool that performs this data integration was recently developed, the integrative Metabolic Analysis Tool (iMAT) [32]. Briefly, taking into account omics data, iMAT predicts flux activity that is stoichiometrically consistent across the metabolic network in a global manner. It determines a subset of reactions to be confidently active or inactive, while leaving pathways that have alternative flux distributions as unknown due to the existence of isozymes or alternative metabolic pathways. By integrating the omics data with a metabolic model encompassing the pertaining biochemical knowledge, iMAT provides a more comprehensive and accurate prediction of flux activity, reflecting the effects of possible post-transcriptional and post-translational regulations that go beyond the information embedded in the raw gene expression data [33]. For example, consider a metabolic pathway composed of three enzymes that are highly active in a given condition. The transcription of only one of the enzymes is up regulated, while the transcription of the other two enzymes remains unchanged. Conventional bioinformatic pathway enrichment analysis based solely on transcriptional data will fail to denote this pathway as highly active, however iMAT analysis could be more successful, if the pathway's activation results in a global network flux distribution that is best consistent with the overall gene expression input (Figure 1).

In this study we applied iMAT analysis to transcriptome data and a genome scale metabolic model of L. monocytogenes to better understand the bacterial metabolic requirements during growth in macrophage cells. Using this approach several metabolic pathways were predicted active or inactive during intracellular growth, potentially reflecting the metabolites availability within these cells. We experimentally deleted key genes in the pathways predicted to be active, validating their contribution to L. monocytogenes intracellular replication and studied the association of the corresponding metabolites in regulation of L. monocytogenes virulence genes.
Results
Conventional transcriptome-based pathway enrichment analysis of metabolic pathway changes during intracellular growth
The global transcriptional profiles of L. monocytogenes (strain 10403S) growing in two different conditions were assessed using whole genome microarray analysis. RNA was extracted from L. monocytogenes grown either inside macrophages (at 6 hours post infection, representing cytosolic replicating bacteria) or in brain heart infusion (BHI) broth (mid log phase). Transcription levels in intracellular bacteria were designated as relative to those in bacteria growing in BHI. Gene expression was coded as 1, −1 or 0 for each gene, indicating a minimum of twofold increase, decrease, or no change, respectively, in transcript levels during intracellular growth versus growth in BHI (Table S1). Next, we retrieved the list of metabolic reactions of L. monocytogenes from the automated genome scale metabolic model reconstruction pipeline [31]. This model consists of 1254 reactions and 816 metabolic genes, encompassing 102 SEED metabolic pathway annotations [34]. Genes in the transcriptome array were assigned to metabolic reactions and their expression levels were converted into reaction activity based on the model's built-in gene-reactions mapping matrix (i.e. enzymatic complexes and isozymes). We then applied a standard hypergeometric test for pathway enrichment taking into account each metabolic reaction observed to change (up or down) to determine which pathways were enriched for altered activities. A metabolic pathway significantly enriched with up-regulated reactions was considered highly active, while a metabolic pathway enriched with down-regulated reactions was considered inactive (Table 1). In agreement with previous studies of metabolic pathways that contribute to L. monocytogenes growth during in vitro infection, we identified arginine biosynthesis, branched chain amino acids (BCAA) biosynthesis and glycerol utilization as pathways highly active during intracellular replication [10], [11], [16]. In addition to the known pathways, our analysis predicted two additional pathways as induced intracellularly: the histidine biosynthesis and L-rhamnose utilization pathways. Along with these highly active pathways, we identified two metabolic pathways to be specifically down regulated during intracellular growth: the formaldehyde assimilation genes and the bacterial fatty acids biosynthesis pathway (FASII).
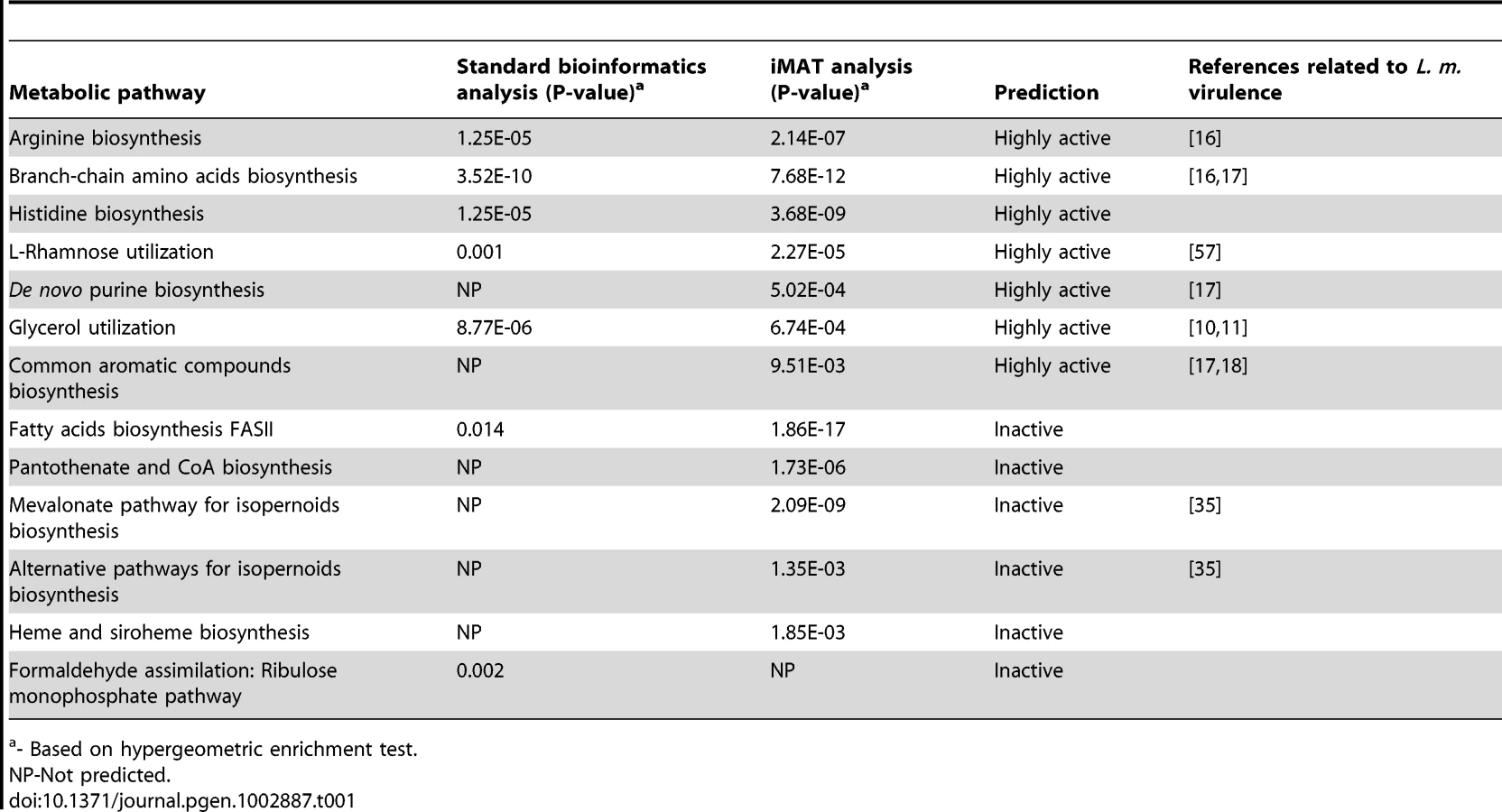
Model-based iMAT analysis of metabolic pathway changes during intracellular growth
Next, we applied the iMAT algorithm to predict metabolic flux activity during intracellular growth of L. monocytogenes. We defined a simulated medium that best represents the cytosolic environment (Table S2), successfully yielding a predicted in silico generation time of 55 min/gen, similar to the one determined experimentally (57 min/gen). iMAT returns a three-valued output, rating the predicted activity of each reaction as confidently active, confidently inactive, or unknown. This output was analyzed by computing the hypergeometric enrichment score of each pathway based on the predicted tri-valued flux activity of each of its reactions (this time determined by iMAT in a global, model based manner) (Table 1). In line with the literature and our conventional gene expression analysis, the arginine, histidine and BCAA biosynthesis pathways together with L-rhamnose and glycerol utilization pathways were predicted to be highly active during intracellular growth. In addition, however, iMAT analysis identified two other metabolic pathways as active intracellularly: the common aromatic compounds biosynthesis (shikimate) pathway and the de novo purine biosynthesis pathway. Notably, the importance of the de novo purine biosynthesis pathway was highlighted by iMAT predictions of metabolites that are imported or exported during intracellular growth (based on the global fluxes and directionality of reactions) (Table S3). Indeed, iMAT predicted uptake of all nucleotides from the host, except for adenosine, explaining the need for de novo purine biosynthesis. Furthermore, iMAT designated five metabolic pathways as inactive/down regulated: the FASII biosynthesis pathway, in line with our conventional gene expression analysis; the mevalonate and the alternative isopernoids biosynthesis pathways [in agreement with [35]]; and two newly identified pathways, the heme biosynthesis and pantothenate-coenzyme A biosynthesis pathways. Statistical analysis of iMAT accuracy versus conventional gene expression analysis, based on a literature database of metabolic pathways that were reported for their role in L. monocytogenes intracellular growth (Table S4), revealed that iMAT predictions are more accurate (P-Value = 0.0001 vs. P-Value = 0.052, hypergeometric distribution test). The precision and recall for iMAT analysis are P = 0.583, R = 0.5, compared to that of the gene expression analysis P = 0.428, R = 0.357. Overall iMAT analysis resulted in a predicted metabolic shift in 12 metabolic pathways upon growth of L. monocytogenes within macrophages, five of which were missed by conventional gene expression analysis. Notably, both methods predicted the novel contribution of histidine biosynthesis and rhamnose utilization to L. monocytogenes intracellular replication.
Experimental validation of the predicted metabolic pathways induction during intracellular growth
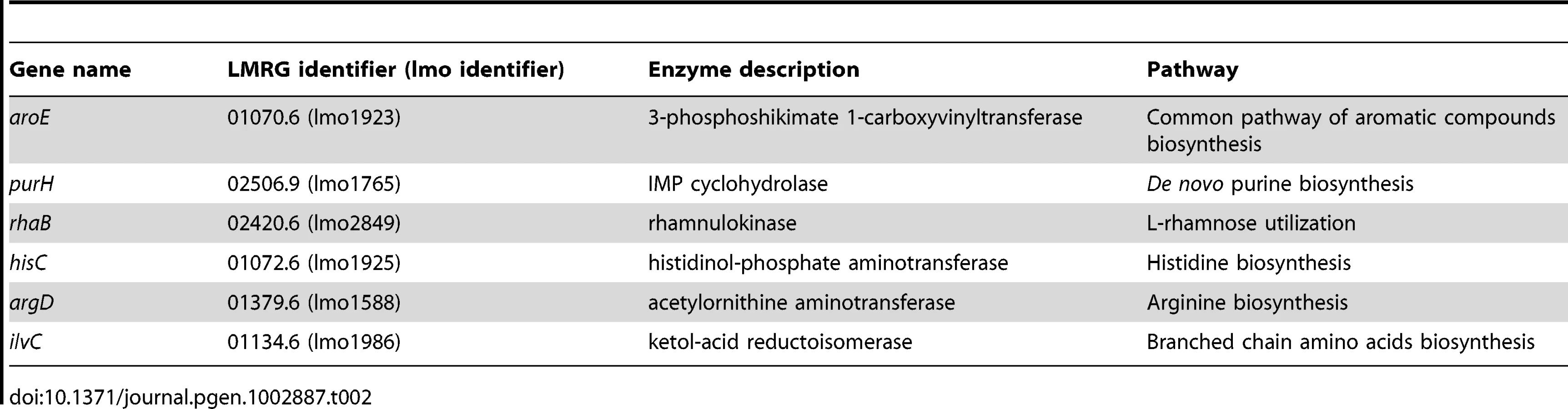
Next, we decided to validate the essentiality of all the metabolic pathways predicted by iMAT analysis to be active during intracellular growth, except for the glycerol utilization pathway that has already been studied in great detail [10], [11]. We chose key listerial enzymes in the active pathways and assessed directly their transcription levels during intracellular growth, as a validation for the microarray data, as well as the consequences of non-polar in - frame deletions of the respective genes. In total, six genes encoding key enzymes from six different metabolic pathways were subjected to further study (Table 2). As depicted by the metabolic map of L. monocytogenes all the chosen genes are central and cannot be compensated (Figure 2A). The genes transcription during intracellular growth of L. monocytogenes in macrophage cells versus growth in BHI medium was assessed using quantitative real-time PCR (RT-qPCR). All six metabolic genes were up-regulated during intracellular growth whereas rpoD and bglA, two control genes, remained unchanged. As another control the transcription level of fabF gene (lmo2201), part of the FASII pathway that was predicted to be down-regulated intracellularly, was analyzed and indeed this gene was shown to be down-regulated during intracellular growth in macrophage cells in comparison to growth in BHI media (Figure 2B). rhaB, ilvC and argD were highly induced, at least 64 fold, while purH, hisC and aroE were induced 2–8 fold (Figure 2B). These enhanced transcriptional levels confirmed that indeed the six pathways are induced and active during L. monocytogenes intracellular growth. Next, we knocked out each one of the selected genes in L. monocytogenes 10403S strain and analyzed the growth capabilities of the resulting six mutants (Table 2). We first tested whether the mutants are indeed defective in their corresponding metabolic pathways. Mutants were grown in minimal defined medium (MDM) [36] and in MDM media that was specifically depleted of the cognate pathway metabolite (Figure S1). As shown in Figure S1, while most mutants grew normally in MDM media (except for ΔaroE and ΔpurH mutants) each one demonstrated a growth defect when the metabolite of their targeted pathway was depleted. As expected, introducing back the corresponding genes complemented the growth defects of these mutants. When tested for growth in BHI, all mutants, with the exception of ΔaroE (as reported also in [18]), grew like WT bacteria, demonstrating that these metabolic pathways are not essential for growth in rich medium (Figure S2). Next, we assessed the contribution of the predicted pathways to intracellular growth of L. monocytogenes. Macrophage cells were infected with the different mutants and intracellular growth was measured. As shown in Figure 3A, all the metabolic mutants replicated less efficiently in macrophage cells in comparison to WT bacteria or to the complemented strains (Figure 3A and Figure S3). The intracellular growth of ΔaroE was highly impaired, however as shown before this gene is also required during growth in rich medium (Figure S2 and [18]). To exclude the possibility that the metabolic mutants are defected in phagosomal escape, we performed a microscope based escape assay, and demonstrated that all mutants were able to escape the macrophages phagosomes like WT bacteria, with the exception of ΔaroE that had a minor defect (Figure 3B). Taken together, these results validate that each of the six predicted pathways is required for intracellular replication of L. monocytogenes, and importantly, establish for the first time a role for histidine biosynthesis and rhamnose catabolism during L. monocytogenes intracellular growth.


Metabolic requirements influence the expression of major virulence genes
Our analyses established that four anabolic pathways are specifically important during L. monocytogenes growth in macrophage cells, namely the BCAA, histidine, arginine and purine biosynthesis pathways. This finding surmises that limited amounts of these nutrients are available in the host cytosol. Given that the abundant availability of non-PTS sugars in the host cytosol is suspected to serve as a signal of intracellular location (see introduction), we investigated if low availability of BCAAs, histidine, arginine and purine might serve similarly as an intracellular signal that modulate the transcription of the virulence genes. Therefore, we tested whether the regulation of these four pathways is linked to the regulation of virulence genes. To this end, WT L. monocytogenes was grown in BHI, minimal defined medium (MDM) and MDM with reduced concentrations (10-fold less) of arginine, histidine, adenine and BCAAs (i.e. isoleucine, leucine and valine), which aimed to mimic the intracellular free amino acids concentration [37]–[39]. The transcription levels of corresponding metabolic genes and virulence genes (prfA the master virulence activator gene, hly gene encoding for LLO, and actA gene) were then analyzed at mid-log phase. The expression of the metabolic genes (purH, ilvC, hisC and argD) was induced during growth in MDM relative to growth in BHI rich medium (Figure 4A), indicating that MDM already contains limiting concentrations of their cognate nutrients. Growth in MDM with ∼10-fold reduced concentrations of arginine, histidine, adenine, isoleucine, leucine and valine resulted in a prolonged lag phase (Figure S4) and in further induction of the BCAA biosynthesis pathway, as shown by increased expression of the ilvC gene (Figure 4A). Remarkably, while growth in MDM resulted in moderate up-regulation of the virulence genes compared to BHI, limiting further the concentrations of arginine, histidine, adenine and BCAAs triggered robust induction of all three virulence genes (Figure 4B). This dramatic response was specific to nutrients synthesized by the intracellularly active metabolic pathways, as growth in MDM with reduced concentrations of other amino acids such as phenylalanine and tryptophan had no effect on expression of the virulence genes (Figure 4B).

Next we tested whether the induction of the virulence genes requires low concentrations of all or some of the identified nutrients. When WT L. monocytogenes was grown in MDM depleted of each nutrient or combinations of nutrients, we found that limited concentrations of BCAAs is the signal for inducing expression of virulence genes (Figure 4C). Among the three BCAAs, isoleucine was found to be the most important for induction of the virulence genes, while leucine and valine presented only a minor contribution (Figure 4D). To explore the dynamic of this response we followed the induction of hly gene (encoding for LLO toxin) during growth in BHI, MDM and MDM with reduced concentration of isoleucine, using a lux reporter system. Wild type bacteria were conjugated with an integrative plasmid containing lux operon under the regulation of the hly promoter (pPL2-Phlylux) [40]. Bacteria were subjected to growth in the different media and parallel measurements of optical density (O.D.600 nm) and luminescence were taken. As shown in Figure 5A, a robust induction of hly promoter was observed during logarithmic phase under limiting concentrations of isoleucine. Notably, this response was not observed during stationary phase in any of the tested conditions, suggesting that a specific regulatory mechanism involves the activation of the virulence genes under low concentrations of isoleucine (Figure 5A–5B). In Gram-positive bacteria, including L. monocytogenes, a nutrient responsive regulator that binds directly isoleucine is known and named CodY. CodY, when bound to isoleucine (and/or to GTP) represses genes that are required for adaptation to nutrient limitation [41]–[44]. Thus, we next examined whether CodY is involved in the regulation of L. monocytogenes virulence genes under low isoleucine concentrations. A ΔcodY complete gene deletion mutant was generated, conjugated with pPL2-Phlylux plasmid and tested under similar conditions. Interestingly, deletion of codY gene did not result in activation of hly transcription, as predicted by its role as a repressor, rather the induction of the hly promoter under isoleucine limiting conditions was strongly dependent on CodY (Figure 5C). In this experiment the hly promoter was activated only in WT bacteria grown in MDM with low concentrations of isoleucine. Similarly, prfA, the master virulence activator gene and actA, the actin-polymerizing gene, were also not induced in the ΔcodY mutant in comparison to WT bacteria. This phenotype was complemented by introducing a copy of codY gene to the ΔcodY mutant, using the chromosomal integrative pLIV2 plasmid (pLIV2-codY; under the control of an IPTG inducible promoter) (Figure 5D).

In light of these findings, we tested whether CodY has a similar role in the regulation of some relevant metabolic genes. To this end, the transcription levels of ilvC and purH genes were analyzed in the ΔcodY mutant and WT bacteria grown under varying concentrations of BCAAs. Notably, we observed that while ilvC and purH were highly induced when BCAAs were limiting (250 and 15-fold induction, respectively), this induction was largely dependent on CodY. However, under high BCAAs concentrations (i.e. in BHI), the ΔcodY mutant also exhibited up-regulation of ilvC and purH, but to a much lesser extent than under conditions of BCAAs depletion alone (50-and 3-fold induction, respectively) (Figure 5E). Overall, these findings indicate that while CodY and BCAAs generally repress the expression of metabolic genes, under BCAA limitation CodY is primarily involved in activation of both metabolic and virulence genes. Finally, analysis of the intracellular growth of the ΔcodY mutant in macrophage cells demonstrated that CodY is required for L. monocytogenes efficient intracellular replication, a phenotype that was complemented by in trans expression of codY gene (pLIV2-codY) (Figure 5F).
CodY plays a positive role in prfA transcription specifically from prfA P1P2 promoters
To study further whether CodY regulation of the virulence genes (i.e., hly and actA genes) is direct or mediated by PrfA, the induction of both hly and prfA promoters by CodY was assessed using the lux reporter system. prfA transcription is initiated from two distinct regions, one proximal encoding two overlapping promoters named P1 and P2, and a second distal region encoding the P3 promoter located up-stream of plcA-prfA genes (Figure 6A) [45]. While the latter requires PrfA itself for co-transcription of plcA-prfA [45], the transcription from P1P2prfA promoters is known to be PrfA-independent (Figure 6A) [46], [47]. Both prfA promoter regions were cloned up-stream to the lux operon in the pPL2-lux plasmid, resulting in pPL2-P1P2prfAlux and pPL2-P3plcA/prfAlux. Next, these plasmids and pPL2-Phlylux were introduced, separately, to ΔprfA, ΔcodY and WT bacteria and promotor activities were measured under BCAAs limiting conditions (i.e., during growth in MDM media with low concentrations of BCAAs). Notably, while induction of hly and P3 promoters required both PrfA and CodY (Figure 6B–6C), the induction of P1P2prfA promoters was dependent solely on the codY gene (Figure 6D). These observations led us to conclude that CodY plays a positive role in the regulation of PrfA expression via the P1P2 promoters, while its role in the regulation of hly and plcA-prfA genes is mediated by PrfA itself. Overall the data presented here clearly demonstrate that CodY is a critical factor controlling PrfA expression linking L. monocytogenes metabolism and virulence.

Discussion
In this study we examined the metabolism of the human bacterial pathogen L. monocytogenes during infection using both a standard bioinformatics pathway enrichment analysis and an integrative computational analysis of transcriptome data using the iMAT tool. This analysis yielded the prediction of metabolic pathways that are highly active and inactive during intracellular growth of L. monocytogenes, inferring the nutrients availability within the cytosolic niche. We validated the contribution of the predicted active pathways to L. monocytogenes growth by generating gene deletion knockouts and monitoring replication rates of these mutants in primary macrophages cells. Importantly, we used this information to search for metabolic requirements that control L. monocytogenes virulence. We found that low concentrations of BCAAs signal the bacteria for its intracellular location and that the isoleucine responsive regulator, CodY, is responsible for the up-regulation of L. monocytogenes virulence genes under these conditions.
The metabolism of L. monocytogenes during intracellular growth differs from its metabolism when growing in rich media. Specifically, this study revealed 12 metabolic pathways to be differentially active during infection. In accordance with previous reports the glycerol utilization pathway was identified as active during intracellular growth [10], [17]. However, it is known that L. monocytogenes can utilize additional carbon sources and most likely switch between them according to cytosolic availability. During residence in the host cytoplasm, L. monocytogenes scavenge for amino acids and vitamins. While most amino acids are acquired from the host [48], we confirmed, and in the case of histidine discovered, that several amino acid biosynthesis pathways are important for bacterial growth during infection. The requirement for arginine biosynthesis by L. monocytogenes likely reflects the low concentrations of this amino acid in host cytosol, as this amino acid is conditionally essential in mammalian cells [49]. In addition, arginine biosynthesis could serve to generate a nitrogen pool, since arginine can be degraded into citrulline and ammonia by arginine deiminase [21], [50]. The requirements for BCAA and histidine biosynthesis could be explained by the fact that these amino acids are not produced by human cells and therefore cytosolic concentrations could be limiting for pathogenic bacteria [49], [51]. Support for this premise comes from studies of another cytosolic pathogen, Burkholderia pseudomalleus, similarly found to require the BCAA and histidine biosynthesis pathways for intracellular growth [52], [53].
We show here that L. monocytogenes require an active histidine biosynthesis pathway for efficient intracellular growth. The HisC enzyme functions also in the synthesis of aromatic amino acids; however these amino acids were shown to be non-essential for L. monocytogenes intracellular replication [18]. Briefly, although the common aromatic compounds biosynthesis pathway was considered initially to be essential for L. monocytogenes intracellular growth, an elegant study by Goebel and colleagues delineated that the growth defect of aro mutants during infection is not due to the lack of aromatic amino acids, but rather to a requirement for menaquinone synthesis. Menaquinone takes part in the respiratory electron chain and this finding explained why the common aromatic compounds biosynthesis pathway is essential during aerobic growth of L. monocytogenes in rich medium but not under anaerobic conditions [18]. Notably, these observations point indirectly to the presence of oxygen in the host cytosol at a level sufficient for aerobic respiration. In accordance, iMAT analysis predicts the uptake of O2 and efflux of CO2 by L. monocytogenes during intracellular growth (Table S3).
The requirement for an active purine (adenine) biosynthesis pathway during L. monocytogenes intracellular growth is in accordance with a similar constraint demonstrated by other intracellular pathogens, such as Brucella melitensis, Mycobacterium tuberculosis, Bacillus anthracis and Burkholderia pseudomallei, and suggests limiting concentrations of purines in mammalian cells [53]–[56]. Of note, iMAT analysis identified the de novo purine biosynthesis pathway as induced during infection whereas conventional gene expression analysis failed to do so, implying of post-transcriptional up-regulation in this pathway. The iMAT prediction was validated as the ΔpurH mutant displayed impaired growth in macrophages.
In addition to the discovery that histidine biosynthesis is required for L. monocytogenes intracellular growth, we show here for the first time that the rhamnose utilization pathway is induced and important during infection. This finding relates to an old observation that rhamnose fermentation could serve as a marker to distinguish between pathogenic and non-pathogenic isolates of L. monocytogenes [57]. As animals do not synthesize rhamnose, it is unlikely that rhamnose utilization provides an alternative carbon source during intracellular bacterial growth. However, rhamnose is an important and abundant carbohydrate in the cell wall of many bacterial species, including L. monocytogenes [58], [59], and accordingly is involved in several cell envelope processes. Indeed, an Escherichia coli ΔrhaB mutant displays higher resistance to cell wall antibiotics [60], supporting the hypothesis that rhamnose catabolism and turnover affect cell wall composition. Notably, a Bacillus anthracis mutant defective in dTDP-rhamnose synthesis was shown to adhere less to macrophages [61] and subsequently, it was reported that the macrophage CD14-MacI complex binds rhamnose residues and promotes bacterial internalization [62]. In this study we established that the L. monocytogenes rhaB gene encoding rhamnulose kinase is transcriptionally up regulated a 100-fold during intracellular growth. In L. monocytogenes rhamnose residues are known to decorate teichoic acids and were suggested to be antigenic determinants [58]. In light of our finding that the rhamnose utilization pathway is induced during infection, it is tempting to speculate that during intracellular growth L. monocytogenes modifies its teichoic acids to evade recognition and avoid triggering the innate immune system. The observed metabolic shift in rhamnose catabolism could directly affect the composition of the cell wall and in this way, bacterial survival during infection. Future studies are necessary to delineate why this pathway is actively required during L. monocytogenes intracellular growth. Overall, our findings that several amino acid biosynthesis pathways are required for growth of L. monocytogenes in macrophages imply that these essential metabolites might be limited in the host cytosol or that the bacteria do not transport them efficiently during intracellular growth. Differentiation between these possibilities requires further studies. It will also be important to discover if these nutrients are limiting in the absence of infecting bacteria or alternatively, are actively depleted from macrophage cytosol as a mechanism to prevent bacterial replication.
Pathogenic bacteria must express virulence factors to exploit nutrients within their host therefore it is not surprising that metabolism and virulence are closely linked. This intimate relationship is best demonstrated by the finding that intracellular carbon sources trigger up regulation of PrfA [12], [63], and thus promote escape into the cytosol and spread to neighboring cells. Here we provide an additional regulatory mechanism connecting intracellular metabolism and virulence, i.e., that low availability of BCAAs, and primarily of isoleucine, leads to the induction of PrfA and in turn, of PrfA-regulated genes. The isoleucine binding transcription regulator, CodY, is directly linked to this response and, as shown, responsible for a positive regulation of prfA. Notably, biosynthesis of BCAAs lies at the crossroads of bacterial metabolism. The synthesis of valine and leucine requires the α-keto acid pyruvate and an amino group from glutamate, and thus is dependent on both glycolysis and the TCA cycle. Furthermore, isoleucine is synthesized from α-ketobutyrate, which is required for sulfur metabolism. Thus BCAA levels indicate the overall carbon, nitrogen, and sulfur metabolic status of the bacteria [64]. In other Gram-positive bacteria CodY was demonstrated to regulate genes involved in diverse processes such as adaptation to starvation, sporulation, biofilm and virulence [43], [65]. Although initial studies indicated a role for CodY as a general repressor (when bound to GTP and/or BCAA), later reports in Bacillus subtilis and Streptococcus pneumonia have demonstrated that it can also function as a transcriptional activator [66]–[68]. Accordingly, in some Gram-positive pathogens the expression of virulence genes was shown to be diminished in CodY null mutants [68]–[72]. In L. monocytogenes, CodY was shown to repress genes involved in amino acid metabolism, nitrogen assimilation and sugar uptake, and was suggested to affect virulence via a functional association with RelA [42]. Our data support the notion that CodY plays a role as repressor of metabolic genes in the presence of BCAAs. However, in this study we show that in the absence or under trace amounts of BCAAs, CodY is important for the activation of virulence genes and metabolic genes necessary for intracellular growth. We demonstrated that CodY specifically affects the expression of prfA via the proximal P1P2 promoters. It is still unclear whether CodY directly interacts with the prfA promoters or that this effect is mediated indirectly by another CodY-regulated protein. In summary, we propose that there are several regulatory connections between L. monocytogenes intracellular metabolism and virulence, with BCAAs and carbon source availability playing a key role in the cytosolic nutrient signature that signals the bacteria for their intracellular location.
This study is the first to address directly the ability of iMAT to serve as a tool to study the metabolic requirements of microorganisms in their natural habitat. We establish that iMAT analysis provides a more comprehensive overview of the activity of the metabolic network of an organism in a given condition. Yet, model-based analyses such as iMAT still suffer from limitations, primarily due to incomplete annotation. In addition, metabolic virulence factors like Hpt and LplA1 [13], [20] can be overlooked, as these are typically not co-regulated with a metabolic pathway. Nevertheless, the approach we presented here can be applied to other bacterial pathogens to reveal novel insights concerning pathogen specific metabolic pathways, and expand our understanding of host-pathogen metabolic interactions. This study highlights the concept that bacterial pathogens have not only acquired dedicated metabolic virulence factors but also changed the regulation of their core metabolism to be able to grow inside their hosts and trigger the virulent response in the right time; further support for the emerging view that pathogen metabolism and virulence are closely interlinked.
Materials and Methods
Ethics statement
The use of animals in this study was limited to preparation of bone marrow derived macrophages from mice. Experimental protocols were approved by the Tel Aviv university Animal Care and Use Committee (L-09-008) according to the Israel Welfare Law (1994) and the National Research Council guide (Guide for the Care and Use of Laboratory Animals 2010).
Bacterial strains and growth media
L. monocytogenes 10403S was the wild type strain (WT) and served as the parental strain when generating gene deletion mutants (Table 2). E. coli XL - 1 Blue (Stratagene) was used for vector propagation and E. coli SM-10 strain [73] was used for plasmid conjugation to L. monocytogenes. The full list of the strains and plasmids used in this study are described in Table S5. Primers used in this study are described in Table S6. L. monocytogenes bacteria were grown at 37°C with agitation in brain heart infusion (BHI) as rich medium or in minimal defined medium (MDM), which is identical to the improved minimal media (IMM) described in [36]. MDM medium includes: phosphate buffer, 0.41 mg/ml magnesium sulfate, 10 mg/ml glucose, 100 µg/ml of each amino acid (i.e., leucine, isoleucine, valine, methionine, arginine, histidine, tryptophan, glutamate, cysteine and phenylalanine), 600 µg/ml glutamine, 0.5 µg/ml biotin, 0.5 µg/ml riboflavin, 20 µg/ml ferric citrate, 1 µg/ml para-aminobenzoic acid, 5 ng/ml lipoic acid, 2.5 µg/ml adenine, 1 µg/ml thiamine, 1 µg/ml pyridoxal, 1 µg/ml calcium pantothenate and 1 µg/ml nicotinamine. For growth under limiting concentrations of nutrients, MDM was freshly made with 10-fold less of the indicated nutrients: BCAAs, histidine, arginine, adenine, phenylalanine and tryptophan (resulting in a final concentration of 10 µg/ml for amino acids and 0.25 µg/ml for adenine).
Bacterial infection of macrophages
Bone marrow derived macrophages (BMDM) were used for infection experiments and were isolated from 6–8 week old female C57/BL6 mice (Harlan laboratories) as described previously [74]. BMDM were cultured in DMEM based media supplemented with 20% fetal bovine serum, sodium pyruvate (1 mM), L-glutamine (2 mM), b-Mercaptoethanol (0.05 mM), and M-CSF (L929-conditioned medium). Approximately 8×106 L. monocytogenes were used to infect 2×106 macrophage cells seeded in a 60 mm Petri dish, resulting in 1–2 bacteria per cell. Thirty minutes after infection, macrophage monolayers were washed three times with PBS and fresh media added. At 1 hour post infection (h.p.i.) gentamicin (50 µg/ml) was added to limit bacterial extracellular growth. Intracellular growth was evaluated as follows. Macrophages were seeded on 13 glass cover slips in a 60 mm plate. At each time point three cover slips were removed and transferred to 2 ml of double-distilled sterile water, which released intracellular bacteria. Then serial dilutions of this 2 ml were plated on BHI plates and colony-forming units (CFUs) counted the next day. Phagosomal escape assay was performed as previously described [75], WT and metabolic mutants of L. monocytogenes expressing GFP (pPL2-GFP integrative plasmid) [65] were used to infect BMDMs on 20 mm slides. Cells were fixed at 2.5 h.p.i. with 4% paraformaldehyde-PBS and permeabilized with Triton X-100. Actin was stained with rhodamine phalloidin (Biotium), and DNA with DAPI containing Vectashield mounting media. Images were taken using Zeiss LSM 510-META confocal microscope.
Bacterial RNA purification and microarray analysis
RNA was harvested from bacteria growing mid-exponentially in BHI medium and from bacteria growing inside macrophages 6 h.p.i. (at which time the bacteria are cytosolic) as described previously [76]. Briefly, bacteria were harvested by filtration and the filters were frozen rapidly in liquid nitrogen. Later, filters were washed and bacterial RNA isolated using phenol-chloroform extraction. Bacterial RNA was amplified using MessageAmp™ II Bacteria Prokaryotic RNA Kit (Ambion). Microarray analysis was performed as described previously [74]. L. monocytogenes microarrays were printed at the Microarray Core Facility of University of California San-Francisco, using the L. monocytogenes oligo set designed and provided by The Institute for Genomic Research (TIGR) [76]. Statistical analysis was performed using the Statistical Analysis of Microarrays (SAM), with the false discovery rate set to 1%. Experiments were performed in duplicates. The values represent two biological repeats (N = 2).
Quantitative real-time PCR analysis
For validation of microarray data the intracellular transcription levels of bacterial metabolic genes were analyzed using real-time quantitative PCR (RT-qPCR) at 6 h.p.i. For the analysis of metabolic genes and virulence genes during growth in BHI medium, MDM and MDM with limiting concentrations of nutrients, RNA was harvested in mid log growth phase at O.D.600 of 0.35. In all cases precultures were grown in MDM media overnight prior to the experiments. One microgram (1 µg) of RNA was reverse transcribed to cDNA using the High Capacity reverse transcription kit (Applied Biosystems). RT-qPCR was performed on 10 ng of cDNA using SYBER Green in a Step-one Plus real time PCR system (Applied Biosystems). The transcription level of each metabolic gene was normalized to that of a reference gene: 16S rRNA in the intracellular experiments and rpoD mRNA in the minimal medium experiments. Statistical analysis was performed using the StepOne™ V2.1 software. RT-qPCR primers are described in Table S6.
Analysis of metabolic pathways (gene expression and iMAT)
For the gene expression analysis, gene expression levels from the microarray analysis were discretized to 1, 0 or −1 for each gene based on a twofold change in expression in intracellular bacteria vs. BHI grown bacteria. The metabolic reactions and pathways of L. monocytogenes were obtained from the genome scale automated metabolic model [31] using PERL scripts. Given the gene expression measurement, the activity state of each enzyme/reaction was determined based on the gene-reaction mapping embedded in the metabolic model, in a standard manner. Pathway based enrichment was computed in a standard bioinformatic manner using a hypergeometric distribution, assigning an enrichment P-value for each pathway for being highly active or inactive. For iMAT analysis, the discretized gene expression levels were incorporated into the metabolic model to predict a set of high and low activity reactions (including post-transcriptional regulation predictions). The iMAT algorithm was described elsewhere [33]. Briefly, a mixed integer linear programming (MILP) problem is solved in order to find a steady-state reaction flux distribution that maximizes the number of reactions whose activity is consistent with their expression state. Subsequently, this list of active/inactive reactions was again analyzed for pathway enrichment in a standard manner, as above. The uptake reactions composing the simulated medium were selected from the pool of available exchange reactions present in the metabolic model, based on a systematic literature search in the Human Metabolome Database. The upper bounds on the set of uptake reactions selected (i.e. allowed import fluxes, measured in mmol h−1 g−1 of dry weight, Table S2) were calibrated based on in silico flux balance analysis simulations of bacterial replication, best representing the experimentally observed bacterial generation time. In both analyses, a correction for multiple hypotheses was done using the false discovery rate (FDR) method. Metabolic pathways with less than 3 reactions were neglected in the analysis. For statistical analysis of predictions accuracy, hypergeometric tests were applied to compare the predictions of both methods to a literature-generated database of metabolic pathways considered essential or non-essential during intracellular growth of L. monocytogenes (Table S4).
Generation of L. monocytogenes gene deletion mutants and complementation strains
All in-frame deletions generated in this work were constructed using L. monocytogenes 10403S strain as the parental strain. Upstream and downstream regions of selected genes were amplified using Phusion DNA polymerase and cloned into pKSV7oriT vector [77]. Cloned plasmids were sequenced and conjugated to L. monocytogenes using E. coli SM-10 strain. L. monocytogenes conjugants were then grown at 41°C for two days on BHI with chloramphenicol to promote plasmid integration into the bacterial chromosome by homologous recombination. For plasmid curing, bacteria were passed several times in fresh BHI without chloramphenicol at 30°C to allow plasmid excision via the generation of an in-frame deletion. Bacteria were then seeded on BHI plates and chloramphenicol sensitive colonies picked for validation of gene deletion by PCR. Complemented strains of deletion mutants were generated by introducing in trans a copy of the deleted gene under the control of its native promoter using the pPL2 integrative vector. For complementation of the codY gene, the integrative pLIV2 plasmid was used, in which codY was cloned under the control of an IPTG inducible promoter [78].
Construction and growth of lux reporter strains
For luminescence assays, a plasmid harboring the lux reporter system was used (pPL2-lux) [40]. For studying the regulation of the hly promoter pPL2-Phlylux was used and was a gift from Colin Hill [40]. For construction of pPL2-P1P2prfAlux and pPL2-P3plcA/prfAlux reporter plasmids, the intragenic regions upstream of the prfA and plcA genes were cloned into the pPL2-lux plasmid. L. monocytogenes precultures were grown in BHI or MDM media (according to experimental conditions) overnight and then diluted to O.D.600 of 0.03 in fresh media as indicated. Bacteria were grown in a Synergy HT Biotek plate reader at 37°C. O.D.600 nm and luminescence measurements were taken every 15 min.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. RayK, MarteynB, SansonettiPJ, TangCM (2009) Life on the inside: the intracellular lifestyle of cytosolic bacteria. Nat Rev Microbiol 7 : 333–340.
2. SwaminathanB, Gerner-SmidtP (2007) The epidemiology of human listeriosis. Microbes Infect 9 : 1236–1243.
3. BierneH, SabetC, PersonnicN, CossartP (2007) Internalins: a complex family of leucine-rich repeat-containing proteins in Listeria monocytogenes. Microbes Infect 9 : 1156–1166.
4. CossartP, VicenteMF, MengaudJ, BaqueroF, Perez-DiazJC, et al. (1989) Listeriolysin O is essential for virulence of Listeria monocytogenes: direct evidence obtained by gene complementation. Infect Immun 57 : 3629–3636.
5. SmithGA, MarquisH, JonesS, JohnstonNC, PortnoyDA, et al. (1995) The two distinct phospholipases C of Listeria monocytogenes have overlapping roles in escape from a vacuole and cell-to-cell spread. Infect Immun 63 : 4231–4237.
6. KathariouS, MetzP, HofH, GoebelW (1987) Tn916-induced mutations in the hemolysin determinant affecting virulence of Listeria monocytogenes. J Bacteriol 169 : 1291–1297.
7. TilneyLG, PortnoyDA (1989) Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes. J Cell Biol 109 : 1597–1608.
8. KocksC, GouinE, TabouretM, BercheP, OhayonH, et al. (1992) L. monocytogenes-induced actin assembly requires the actA gene product, a surface protein. Cell 68 : 521–531.
9. DussurgetO, Pizarro-CerdaJ, CossartP (2004) Molecular determinants of Listeria monocytogenes virulence. Annu Rev Microbiol 58 : 587–610.
10. EylertE, ScharJ, MertinsS, StollR, BacherA, et al. (2008) Carbon metabolism of Listeria monocytogenes growing inside macrophages. Mol Microbiol 69 : 1008–1017.
11. JosephB, MertinsS, StollR, ScharJ, UmeshaKR, et al. (2008) Glycerol metabolism and PrfA activity in Listeria monocytogenes. J Bacteriol 190 : 5412–5430.
12. RipioMT, BrehmK, LaraM, SuarezM, Vazquez-BolandJA (1997) Glucose-1-phosphate utilization by Listeria monocytogenes is PrfA dependent and coordinately expressed with virulence factors. J Bacteriol 179 : 7174–7180.
13. Chico-CaleroI, SuarezM, Gonzalez-ZornB, ScorttiM, SlaghuisJ, et al. (2002) Hpt, a bacterial homolog of the microsomal glucose - 6-phosphate translocase, mediates rapid intracellular proliferation in Listeria. Proc Natl Acad Sci U S A 99 : 431–436.
14. EisenreichW, DandekarT, HeesemannJ, GoebelW (2010) Carbon metabolism of intracellular bacterial pathogens and possible links to virulence. Nat Rev Microbiol 8 : 401–412.
15. FreitagNE, PortGC, MinerMD (2009) Listeria monocytogenes - from saprophyte to intracellular pathogen. Nat Rev Microbiol 7 : 623–628.
16. JosephB, PrzybillaK, StuhlerC, SchauerK, SlaghuisJ, et al. (2006) Identification of Listeria monocytogenes genes contributing to intracellular replication by expression profiling and mutant screening. J Bacteriol 188 : 556–568.
17. SchauerK, GeginatG, LiangC, GoebelW, DandekarT, et al. (2010) Deciphering the intracellular metabolism of Listeria monocytogenes by mutant screening and modelling. BMC Genomics 11 : 573.
18. StritzkerJ, JandaJ, SchoenC, TauppM, PilgrimS, et al. (2004) Growth, virulence, and immunogenicity of Listeria monocytogenes aro mutants. Infect Immun 72 : 5622–5629.
19. SchauerK, StolzJ, SchererS, FuchsTM (2009) Both thiamine uptake and biosynthesis of thiamine precursors are required for intracellular replication of Listeria monocytogenes. J Bacteriol 191 : 2218–2227.
20. KeeneyKM, StuckeyJA, O'RiordanMX (2007) LplA1-dependent utilization of host lipoyl peptides enables Listeria cytosolic growth and virulence. Mol Microbiol 66 : 758–770.
21. JosephB, GoebelW (2007) Life of Listeria monocytogenes in the host cells' cytosol. Microbes Infect 9 : 1188–1195.
22. GarsinDA (2010) Ethanolamine utilization in bacterial pathogens: roles and regulation. Nat Rev Microbiol 8 : 290–295.
23. BuchrieserC, RusniokC, KunstF, CossartP, GlaserP (2003) Comparison of the genome sequences of Listeria monocytogenes and Listeria innocua: clues for evolution and pathogenicity. FEMS Immunol Med Microbiol 35 : 207–213.
24. OberhardtMA, PalssonBO, PapinJA (2009) Applications of genome-scale metabolic reconstructions. Mol Syst Biol 5 : 320.
25. PriceND, ReedJL, PalssonBO (2004) Genome-scale models of microbial cells: evaluating the consequences of constraints. Nat Rev Microbiol 2 : 886–897.
26. KimHU, KimSY, JeongH, KimTY, KimJJ, et al. (2011) Integrative genome-scale metabolic analysis of Vibrio vulnificus for drug targeting and discovery. Mol Syst Biol 7 : 460.
27. PlataG, HsiaoTL, OlszewskiKL, LlinasM, VitkupD (2010) Reconstruction and flux-balance analysis of the Plasmodium falciparum metabolic network. Mol Syst Biol 6 : 408.
28. HuthmacherC, HoppeA, BulikS, HolzhutterHG (2010) Antimalarial drug targets in Plasmodium falciparum predicted by stage-specific metabolic network analysis. BMC Syst Biol 4 : 120.
29. BordbarA, LewisNE, SchellenbergerJ, PalssonBO, JamshidiN (2010) Insight into human alveolar macrophage and M. tuberculosis interactions via metabolic reconstructions. Mol Syst Biol 6 : 422.
30. NavidA, AlmaasE (2009) Genome-scale reconstruction of the metabolic network in Yersinia pestis, strain 91001. Mol Biosyst 5 : 368–375.
31. HenryCS, DeJonghM, BestAA, FrybargerPM, LinsayB, et al. (2010) High-throughput generation, optimization and analysis of genome-scale metabolic models. Nat Biotechnol 28 : 977–982.
32. ZurH, RuppinE, ShlomiT (2010) iMAT: an integrative metabolic analysis tool. Bioinformatics 26 : 3140–3142.
33. ShlomiT, CabiliMN, HerrgardMJ, PalssonBO, RuppinE (2008) Network-based prediction of human tissue-specific metabolism. Nat Biotechnol 26 : 1003–1010.
34. AzizRK, BartelsD, BestAA, DeJonghM, DiszT, et al. (2008) The RAST Server: rapid annotations using subsystems technology. BMC Genomics 9 : 75.
35. BegleyM, BronPA, HeustonS, CaseyPG, EnglertN, et al. (2008) Analysis of the isoprenoid biosynthesis pathways in Listeria monocytogenes reveals a role for the alternative 2-C-methyl-D-erythritol 4-phosphate pathway in murine infection. Infect Immun 76 : 5392–5401.
36. Phan-ThanhL, GormonT (1997) A chemically defined minimal medium for the optimal culture of Listeria. Int J Food Microbiol 35 : 91–95.
37. MartinRE, KirkK (2007) Transport of the essential nutrient isoleucine in human erythrocytes infected with the malaria parasite Plasmodium falciparum. Blood 109 : 2217–2224.
38. KohnoH, KandaS, KannoT (1986) Immunoaffinity purification and characterization of leucine aminopeptidase from human liver. J Biol Chem 261 : 10744–10748.
39. EagleH, PiezKA, LevyM (1961) The intracellular amino acid concentrations required for protein synthesis in cultured human cells. J Biol Chem 236 : 2039–2042.
40. BronPA, MonkIR, CorrSC, HillC, GahanCG (2006) Novel luciferase reporter system for in vitro and organ-specific monitoring of differential gene expression in Listeria monocytogenes. Appl Environ Microbiol 72 : 2876–2884.
41. ShiversRP, SonensheinAL (2004) Activation of the Bacillus subtilis global regulator CodY by direct interaction with branched-chain amino acids. Mol Microbiol 53 : 599–611.
42. BennettHJ, PearceDM, GlennS, TaylorCM, KuhnM, et al. (2007) Characterization of relA and codY mutants of Listeria monocytogenes: identification of the CodY regulon and its role in virulence. Mol Microbiol 63 : 1453–1467.
43. StenzL, FrancoisP, WhitesonK, WolzC, LinderP, et al. (2011) The CodY pleiotropic repressor controls virulence in gram-positive pathogens. FEMS Immunol Med Microbiol 62 : 123–139.
44. VillapakkamAC, HandkeLD, BelitskyBR, LevdikovVM, WilkinsonAJ, et al. (2009) Genetic and biochemical analysis of the interaction of Bacillus subtilis CodY with branched-chain amino acids. J Bacteriol 191 : 6865–6876.
45. FreitagNE, PortnoyDA (1994) Dual promoters of the Listeria monocytogenes prfA transcriptional activator appear essential in vitro but are redundant in vivo. Mol Microbiol 12 : 845–853.
46. RauchM, LuoQ, Muller-AltrockS, GoebelW (2005) SigB-dependent in vitro transcription of prfA and some newly identified genes of Listeria monocytogenes whose expression is affected by PrfA in vivo. J Bacteriol 187 : 800–804.
47. OllingerJ, BowenB, WiedmannM, BoorKJ, BergholzTM (2009) Listeria monocytogenes sigmaB modulates PrfA-mediated virulence factor expression. Infect Immun 77 : 2113–2124.
48. EisenreichW, SlaghuisJ, LaupitzR, BussemerJ, StritzkerJ, et al. (2006) 13C isotopologue perturbation studies of Listeria monocytogenes carbon metabolism and its modulation by the virulence regulator PrfA. Proc Natl Acad Sci U S A 103 : 2040–2045.
49. ReedsPJ (2000) Dispensable and indispensable amino acids for humans. J Nutr 130 : 1835S–1840S.
50. RyanS, BegleyM, GahanCG, HillC (2009) Molecular characterization of the arginine deiminase system in Listeria monocytogenes: regulation and role in acid tolerance. Environ Microbiol 11 : 432–445.
51. J D KoppleMES (1975) Evidence that histidine is an essential amino acid in normal and chronically uremic man. J Clin Invest 55 : 881–891.
52. AtkinsT, PriorRG, MackK, RussellP, NelsonM, et al. (2002) A mutant of Burkholderia pseudomallei, auxotrophic in the branched chain amino acid biosynthetic pathway, is attenuated and protective in a murine model of melioidosis. Infect Immun 70 : 5290–5294.
53. PilatzS, BreitbachK, HeinN, FehlhaberB, SchulzeJ, et al. (2006) Identification of Burkholderia pseudomallei genes required for the intracellular life cycle and in vivo virulence. Infect Immun 74 : 3576–3586.
54. DrazekES, HoungHS, CrawfordRM, HadfieldTL, HooverDL, et al. (1995) Deletion of purE attenuates Brucella melitensis 16 M for growth in human monocyte-derived macrophages. Infect Immun 63 : 3297–3301.
55. JenkinsA, CoteC, TwenhafelN, MerkelT, BozueJ, et al. (2011) Role of purine biosynthesis in Bacillus anthracis pathogenesis and virulence. Infect Immun 79 : 153–166.
56. JacksonM, PhalenSW, LagranderieM, EnsergueixD, ChavarotP, et al. (1999) Persistence and protective efficacy of a Mycobacterium tuberculosis auxotroph vaccine. Infect Immun 67 : 2867–2873.
57. GrovesRD, WelshimerHJ (1977) Separation of pathogenic from apathogenic Listeria monocytogenes by three in vitro reactions. J Clin Microbiol 5 : 559–563.
58. KamisangoK, FujiiH, OkumuraH, SaikiI, ArakiY, et al. (1983) Structural and immunochemical studies of teichoic acid of Listeria monocytogenes. J Biochem 93 : 1401–1409.
59. EugsterMRLM (2011) Rapid Analysis of Listeria monocytogenes Cell Wall Teichoic Acid Carbohydrates by ESI-MS/MS. PLoS ONE 6 doi:10.1371/journal.pone.0021500.
60. DuoM, HouS, RenD (2008) Identifying Escherichia coli genes involved in intrinsic multidrug resistance. Appl Microbiol Biotechnol 81 : 731–741.
61. BozueJA, ParthasarathyN, PhillipsLR, CoteCK, FellowsPF, et al. (2005) Construction of a rhamnose mutation in Bacillus anthracis affects adherence to macrophages but not virulence in guinea pigs. Microb Pathog 38 : 1–12.
62. OlivaC, TurnboughCLJr, KearneyJF (2009) CD14-Mac-1 interactions in Bacillus anthracis spore internalization by macrophages. Proc Natl Acad Sci U S A 106 : 13957–13962.
63. StollR, MertinsS, JosephB, Muller-AltrockS, GoebelW (2008) Modulation of PrfA activity in Listeria monocytogenes upon growth in different culture media. Microbiology 154 : 3856–3876.
64. SomervilleGA, ProctorRA (2009) At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci. Microbiol Mol Biol Rev 73 : 233–248.
65. MolleV, NakauraY, ShiversRP, YamaguchiH, LosickR, et al. (2003) Additional targets of the Bacillus subtilis global regulator CodY identified by chromatin immunoprecipitation and genome-wide transcript analysis. J Bacteriol 185 : 1911–1922.
66. ShiversRP, DineenSS, SonensheinAL (2006) Positive regulation of Bacillus subtilis ackA by CodY and CcpA: establishing a potential hierarchy in carbon flow. Mol Microbiol 62 : 811–822.
67. PreisH, EckartRA, GudipatiRK, HeidrichN, BrantlS (2009) CodY activates transcription of a small RNA in Bacillus subtilis. J Bacteriol 191 : 5446–5457.
68. HendriksenWT, BootsmaHJ, EstevaoS, HoogenboezemT, de JongA, et al. (2008) CodY of Streptococcus pneumoniae: link between nutritional gene regulation and colonization. J Bacteriol 190 : 590–601.
69. DineenSS, McBrideSM, SonensheinAL (2010) Integration of metabolism and virulence by Clostridium difficile CodY. J Bacteriol 192 : 5350–5362.
70. van SchaikW, ChateauA, DilliesMA, CoppeeJY, SonensheinAL, et al. (2009) The global regulator CodY regulates toxin gene expression in Bacillus anthracis and is required for full virulence. Infect Immun 77 : 4437–4445.
71. ChateauA, van SchaikW, SixA, AucherW, FouetA (2011) CodY regulation is required for full virulence and heme iron acquisition in Bacillus anthracis. FASEB J 25 : 4445–4456.
72. KrethJ, ChenZ, FerrettiJ, MalkeH (2011) Counteractive balancing of transcriptome expression involving CodY and CovRS in Streptococcus pyogenes. J Bacteriol 193 : 4153–4165.
73. SimonR, PrieferU, PühlerA (1983) A broad host range mobilization system for in vitro genetic engineering: transposon mutagenesis in Gram negative bacteria. Bio/Technology 1 : 784–791.
74. HerskovitsAA, AuerbuchV, PortnoyDA (2007) Bacterial ligands generated in a phagosome are targets of the cytosolic innate immune system. PLoS Pathog 3: e51 doi:10.1371/journal.ppat.0030051.
75. GlomskiIJ, GeddeMM, TsangAW, SwansonJA, PortnoyDA (2002) The Listeria monocytogenes hemolysin has an acidic pH optimum to compartmentalize activity and prevent damage to infected host cells. J Cell Biol 156 : 1029–1038.
76. LeberJH, CrimminsGT, RaghavanS, Meyer-MorseNP, CoxJS, et al. (2008) Distinct TLR - and NLR-mediated transcriptional responses to an intracellular pathogen. PLoS Pathog 4: e6 doi:10.1371/journal.ppat.0040006.
77. LauerP, HansonB, LemmensEE, LiuW, LuckettWS, et al. (2008) Constitutive Activation of the PrfA regulon enhances the potency of vaccines based on live-attenuated and killed but metabolically active Listeria monocytogenes strains. Infect Immun 76 : 3742–3753.
78. LauerP, ChowMY, LoessnerMJ, PortnoyDA, CalendarR (2002) Construction, characterization, and use of two Listeria monocytogenes site-specific phage integration vectors. J Bacteriol 184 : 4177–4186.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 9
Nejčtenější v tomto čísle
- Enrichment of HP1a on Drosophila Chromosome 4 Genes Creates an Alternate Chromatin Structure Critical for Regulation in this Heterochromatic Domain
- Normal DNA Methylation Dynamics in DICER1-Deficient Mouse Embryonic Stem Cells
- The NDR Kinase Scaffold HYM1/MO25 Is Essential for MAK2 MAP Kinase Signaling in
- Functional Variants in and Involved in Activation of the NF-κB Pathway Are Associated with Rheumatoid Arthritis in Japanese