
Regulates Refractive Error and Myopia Development in Mice and Humans
Gene variants identified by GWAS studies to date explain only a small fraction of myopia cases because myopia represents a complex disorder thought to be controlled by dozens or even hundreds of genes. The majority of genetic variants underlying myopia seems to be of small effect and/or low frequency, which makes them difficult to identify using classical genetic approaches, such as GWAS, alone. Here, we combined gene expression profiling in a monkey model of myopia, human GWAS, and a gene-targeted mouse model of myopia to identify one of the “missing” myopia genes, APLP2. We found that a low-frequency risk allele of APLP2 confers susceptibility to myopia only in children exposed to large amounts of daily reading, thus, providing an experimental example of the long-hypothesized gene-environment interaction between nearwork and genes underlying myopia. Functional analysis of APLP2 using an APLP2 knockout mouse model confirmed functional significance of APLP2 in refractive development and implicated a potential role of synaptic transmission at the level of glycinergic amacrine cells of the retina for the development of myopia. Furthermore, mouse studies revealed that lack of Aplp2 has a dose-dependent suppressive effect on susceptibility to form-deprivation myopia, providing a potential gene-specific target for therapeutic intervention to treat myopia.
Published in the journal:
. PLoS Genet 11(8): e32767. doi:10.1371/journal.pgen.1005432
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1005432
Summary
Gene variants identified by GWAS studies to date explain only a small fraction of myopia cases because myopia represents a complex disorder thought to be controlled by dozens or even hundreds of genes. The majority of genetic variants underlying myopia seems to be of small effect and/or low frequency, which makes them difficult to identify using classical genetic approaches, such as GWAS, alone. Here, we combined gene expression profiling in a monkey model of myopia, human GWAS, and a gene-targeted mouse model of myopia to identify one of the “missing” myopia genes, APLP2. We found that a low-frequency risk allele of APLP2 confers susceptibility to myopia only in children exposed to large amounts of daily reading, thus, providing an experimental example of the long-hypothesized gene-environment interaction between nearwork and genes underlying myopia. Functional analysis of APLP2 using an APLP2 knockout mouse model confirmed functional significance of APLP2 in refractive development and implicated a potential role of synaptic transmission at the level of glycinergic amacrine cells of the retina for the development of myopia. Furthermore, mouse studies revealed that lack of Aplp2 has a dose-dependent suppressive effect on susceptibility to form-deprivation myopia, providing a potential gene-specific target for therapeutic intervention to treat myopia.
Introduction
Postnatal refractive eye development is a tightly coordinated process whereby visual experience fine-tunes a genetic program of ocular growth towards an optimal match between the optical power of the eye and its axial length in a process called “emmetropization” [1,2]. The emmetropization process is regulated by a vision-driven feedback loop in the retina and downstream signaling cascades in other ocular tissues, and normally results in sharp vision (emmetropia). Failure to achieve or maintain emmetropia leads to the development of refractive errors, i.e., farsightedness (hyperopia) or nearsightedness (myopia). Myopia is the most common vision disorder worldwide [3]. The prevalence of myopia has increased from 25% to 44% of the adult population in the United States in the last 30 years [4], and reached more than 80% of young adults in some parts of Asia [5,6]. Myopia negatively affects self-perception, job/activity choices, and ocular health [7–9]. Epidemiological data suggest that common myopia represents a major risk factor for a number of potentially blinding ocular diseases such as cataract, glaucoma, retinal detachment, and myopic maculopathy, which is comparable to the risks associated with hypertension for stroke and myocardial infarction, and represents one of the leading causes of blindness [10–12]. It is estimated that 2.5 billion people (1/3 of the world’s population) will be affected by myopia by 2020 [13]. Uncorrected refractive errors are the major cause of vision loss and refractive errors and is one of five World Health Organization’s designated priority health conditions [3,13].
Refractive eye development is controlled by both environmental and genetic factors [14–17]. However, genetic factors are believed to play a key role in determining the impact of environmental factors on refractive eye development, including populations that have experienced rapid rises in the prevalence of myopia in recent decades [14–18]. Human population studies suggest that contribution of genetic factors accounts for 60%-90% of variance in refraction [19–24]. Human genetic mapping studies have identified over 24 chromosomal loci linked to myopia [15,25–30]. However, the currently-identified variants account for only a small fraction of myopia cases [31] suggesting the existence of a large number of yet unidentified low-frequency or small-effect variants, which underlie the majority of myopia cases [32–35]. Here, we present genetic and functional evidence identifying amyloid beta (A4) precursor-like protein 2 (APLP2), which was previously found to be involved in synaptic plasticity and transmission in the central nervous system [36–52], as one such myopia-susceptibility gene.
Results
Genetic variation at the APLP2 locus is associated with myopia in children and adults
In a previous study designed to identify genes differentially expressed in myopic eyes, we performed large-scale gene expression profiling in the retina of green monkeys (Chlorocebus aethiops) with experimentally induced myopia and identified 119 differentially expressed genes [53]. Here, gene set enrichment analysis (GSEA) [54,55] of these data revealed that expression of one of these genes, APLP2, among others was strongly associated with the refractive error phenotype. APLP2 was found to be overexpressed in myopia and suppressed in hyperopia (Fig 1 and S1 Table).
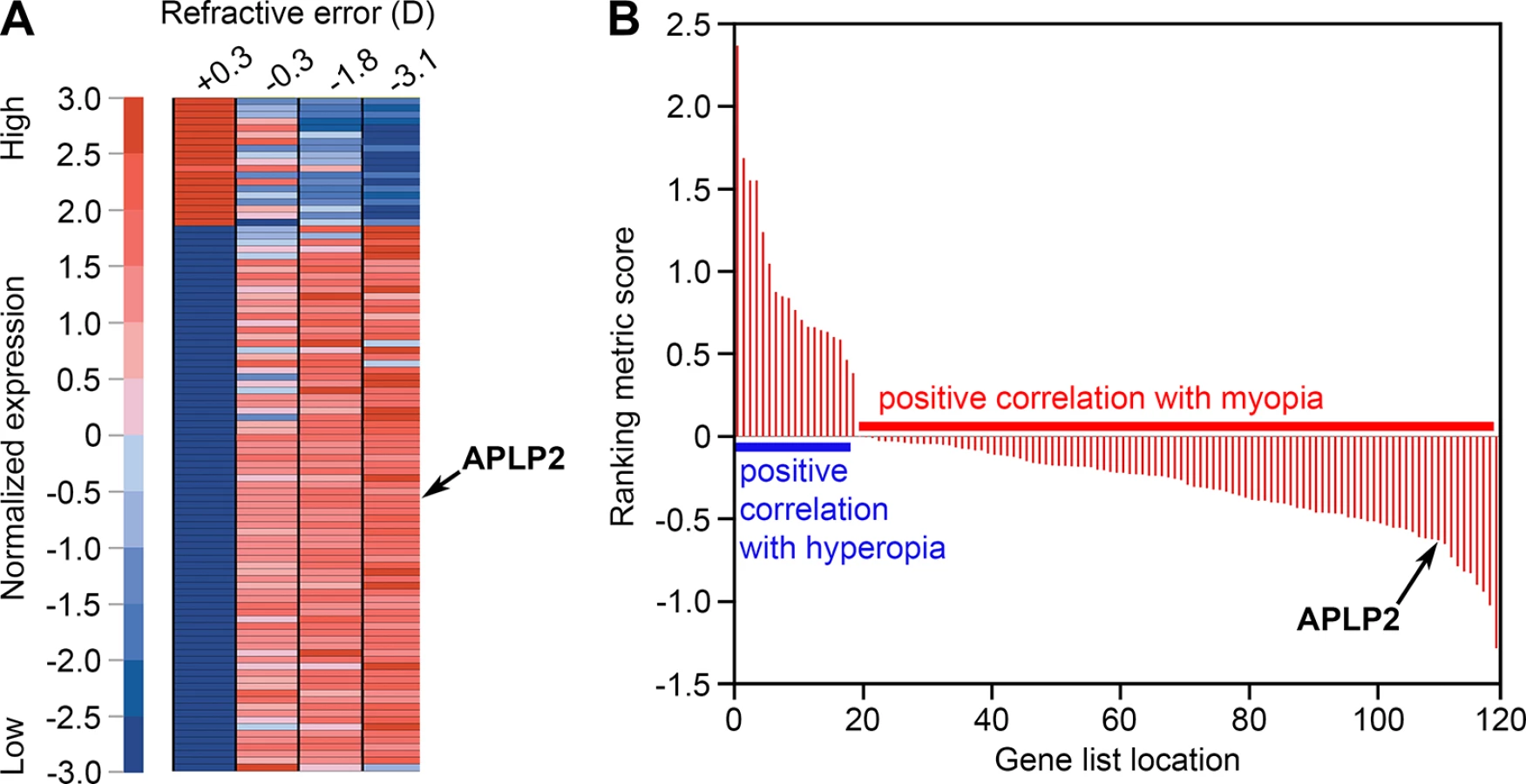
To explore whether genetic variants within or nearby APLP2 influence refractive error development in humans, single nucleotide polymorphism (SNP) genetic variants within 100 kb of the APLP2 gene were tested for association with refractive error in children participating in a UK birth cohort study (the Avon Longitudinal Study of Parents and Children, ALSPAC). Numerous SNPs in an LD block that encompassed the promoter region and 5’-end of the APLP2 gene were associated with refractive error at age 15 years in ALSPAC participants (Fig 2A and 2B). The most strongly associated variant was rs188663068 (risk allele frequency (RAF) = 0.01, n = 3,819, p = 5.0 × 10−4), each copy of the risk allele being associated with a -0.6 D shift in refractive error. Because SNPs in LD do not offer independent evidence of association, permutation testing was used to evaluate whether these results were likely to have arisen by chance. Consistent with the QQ-plot for the full set of SNPs tested (Fig 2B) permutation-based analysis suggested that obtaining a p-value as low as p = 5.0 × 10−4 for rs188663068 was not unexpected; however, such an excess of low p-values was unlikely to have occurred by chance (p = 1.4 × 10−2). These findings are consistent with the notion that an excess of genetic variants in the promoter region and 5’-end of APLP2 are associated with refractive error, but because the associated variants have a low minor allele frequency, no single SNP provides compelling evidence on its own. SNPs within 100 kb of the APLP2 gene were also evaluated in a meta-analyzed refractive error genome-wide association study (GWAS) dataset from the international Consortium for Refractive Error and Myopia (CREAM), which included 45,756 adult individuals from 27 Caucasian and 5 Asian cohorts [30]. As in the ALSPAC cohort, SNPs in the LD block encompassing the promoter region and 5’-end of the APLP2 gene were most strongly associated with refractive error in the CREAM consortium sample (Fig 2C and 2D; top SNP, rs7127037, p = 6.6 × 10−3). The use of permutation testing to account for multiple testing was not possible for the CREAM dataset since we did not have access to the raw genotypes of individual participants. Therefore, as an alternative approach to test for an excess of low p-values in the region found to be associated in the ALSPAC cohort (hg19 chr 11 : 129904497–129971498), the distribution of p-values inside this region was compared to the surrounding region encompassing 100 kb on the either side of the gene. P-values inside the region were skewed towards low values compared with the p-values outside the region (p = 5.0 × 10−3, two-sample Kolmogorov-Smirnov test, S1 Fig).

Gene-environment interaction between APLP2 and time spent reading in children with myopia
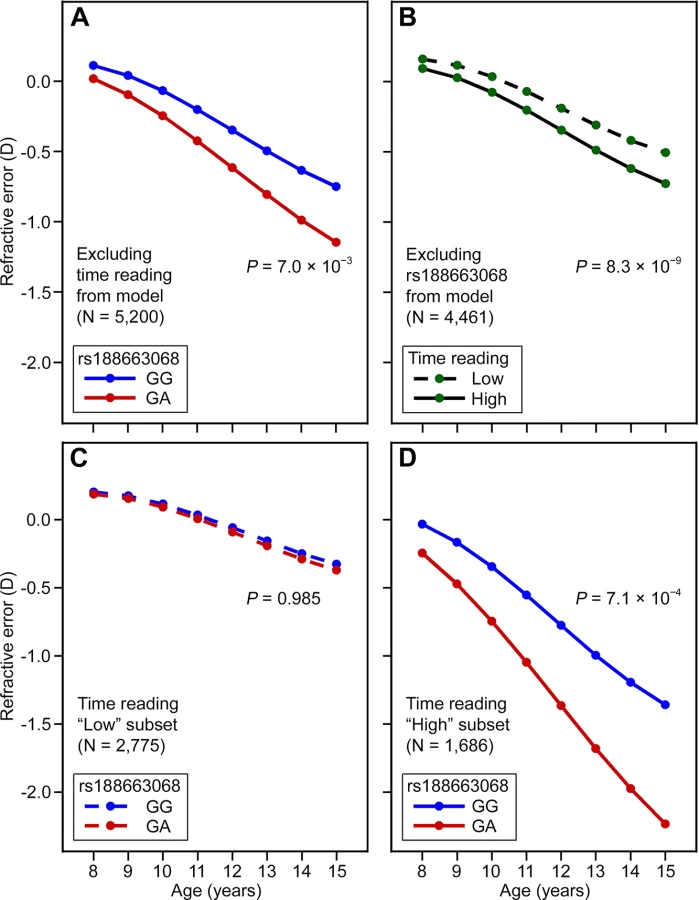
To explore the possibility of an interaction between APLP2 gene variants and visual experience, we exploited the availability of longitudinal refractive error measurements over childhood (age range 8 to 15 years) and prospective exposure information regarding the two most important currently known environmental risk factors for myopia, i.e., time spent reading and time spent outdoors. For the strongest APLP2 risk variant, rs188663068, a “growth trajectory” analysis of refractive development revealed a progressive, age-dependent shift towards a relatively more myopic refractive error in individuals carrying a single copy of the high-risk “A” allele compared to individuals homozygous for the low-risk “G” allele (p = 7.0 × 10−3; Fig 3A and S2 Table). When analysed separately, time spent reading ascertained at age 8–9 years, and categorized as “high” or “low”, was also predictive of refractive trajectory in ALSPAC participants, with the “high” reading group also gradually diverging towards a relatively more myopic refractive error as they became older (p = 8.3 × 10-9; Fig 3B and S3 Table). A model, which included both rs188663068 and time spent reading as predictors, provided strong evidence for a 3-way interaction between age, time spent reading at age 8–9 years and SNP genotype; implicating gene-environment interaction between the APLP2 genetic variant and time spent reading that became greater with age (3-way interaction term, p = 4.0 × 10−3; S4 Table). Stratifying by time spent reading (“low” versus “high”) revealed that the high-risk “A” allele of rs188663068 was predictive of progression towards myopia only in children who spent a “high” amount of time reading (genotype x age interaction term, p = 0.99 and p = 7.1 × 10−4 in “low” and “high” readers respectively; Fig 3C and 3D, S5 and S6 Tables). Logistic regression analysis in the ALSPAC participants at age 15 years confirmed the clinical relevance of the association at the APLP2 locus (S7 and S8 Tables). The odds ratio (OR) for myopia associated with a single copy of the rs188663068 risk allele was 1.98 (95% CI = 1.02 to 3.87, p = 4.5 × 10−2), while by comparison the OR associated with a “high” amount of time spent reading at age 8 years was 1.61 (95% CI = 1.33 to 1.93, p = 6.4 × 10−7). Inclusion of an interaction term between rs188663068 genotype and time spent reading supported the presence of an interaction; the OR for myopia of participants in the “high” reading group and carrying a copy of the risk allele was 5.42 (95% CI = 1.15 to 25.52, p = 3.3 × 10−2). In linear regression analysis, time spent reading, alone, predicted 0.6% of the variance in refractive error at age 15 years, while the percentage of explained variance increased to 0.9% with the inclusion of rs188663068 genotype and a SNP/reading interaction term (S9 and S10 Tables). There was no evidence for an interaction between rs188663068 genotype and time spent outdoors (S11 and S12 Tables).
Aplp2 regulates refractive eye development and susceptibility to myopia in mice
To examine whether APLP2 is functionally involved in refractive error development, we studied refractive eye development in Aplp2 knockout mice (Fig 4). Mice homozygous for a null allele of the Aplp2 gene (Aplp2-/- mice) were found to develop high degrees of hyperopia (+11.5 ± 2.2 D, p < 1.0 × 10−4) compared to both heterozygous (Aplp2+/-) (-0.8 ± 2.0 D, p < 1.0 × 10−4) and wild-type (Aplp2+/+) (+0.3 ± 2.2 D, p < 1.0 × 10−4) littermates (Fig 4A), consistent with the finding that APLP2 expression is suppressed in hyperopia in monkeys (Fig 1). Visual form deprivation induced -1.2 ± 0.6 D of myopia (p = 3.0 × 10−2) in Aplp2-/- mice compared to -5.7 ± 1.1 D (p < 1.0 × 10−4) in Aplp2+/- heterozygotes and -11.0 ± 1.7 D (p < 1.0 × 10−4) in wild-type littermates, indicating that lack of Aplp2 expression has a dose-dependent inhibitory effect on susceptibility to environmentally induced myopia (F(2, 33) = 191.0, p < 1.0 × 10−4) (Fig 4B), thus confirming gene-environment interaction between APLP2 and visual experience identified by human studies.

Aplp2 regulates refractive eye development by modulating the function of glycinergic amacrine cells of the retina
Visual acuity in Aplp2-/- mice was not different from that in heterozygous and wild-type littermates (F(2, 20) = 0.6, p = 0.58) (Fig 4C and S13 Table), whereas contrast sensitivity was reduced compared to both heterozygous and wild-type mice (F(12, 120) = 3.6, p = 1.5 × 10−4) (Fig 4D and S13 Table). Analysis of scotopic ERGs in Aplp2 knockouts revealed that lack of Aplp2 caused a dose-dependent decrease in the amplitude of the b-wave (F(2, 18) = 6.9, p = 6.0 × 10−3) (Fig 5A and 5B and 5D) and oscillatory potentials (F(2, 18) = 3.6–20.5, p < 1.0 × 10−3) (Fig 5F and 5G); as well as an increase in the implicit time of the b-wave (F(2, 18) = 6.1, p = 9.6 × 10−3) (Fig 5C and 5E) and oscillatory potentials (F(2, 18) = 4.5–20.9, p < 5.0 × 10−3) (Fig 5F and 5H). Considering that oscillatory potentials are primarily generated by retinal amacrine cells [56,57], which also modulate the amplitude of the b-wave generated by the bipolar cells [58–66], the ERG data suggested that Aplp2 modulates the function of amacrine cells. Therefore, we then analyzed the expression of Aplp2 in the retina at the mRNA and protein levels. In situ hybridization and immunohistochemical analysis confirmed Aplp2 expression in both bipolar and amacrine cells of wild-type mice (Fig 6A–6C and S2 Fig). Further analysis also revealed that Aplp2 was expressed in glycinergic amacrine cells, but was not expressed in GABAergic amacrines (Figs 6D and 6E and S2).


Discussion
More than 24 chromosomal loci associated with human myopia have been identified [15,25,26,29–31,67–89] either by linkage analysis, which primarily focuses on rare variants with large effect size causing Mendelian forms of myopia, or by large-scale GWAS studies, targeting common variants with moderate effect sizes underlying common myopia. However, refractive error is inherited as a complex quantitative trait thought to be influenced by multiple interacting genes and controlled by dozens and even hundreds of chromosomal loci [19,90–92]. The variants identified to date account for less than 10% of common myopia cases [31], suggesting the existence of a large number of yet unidentified low-frequency and/or small-effect variants, which underlie the majority of myopia cases [32–35].
Several approaches for finding the “missing heritability” of complex traits have been proposed (e.g., increasing GWAS sample sizes, using larger catalogues of human variation, including copy number variations in analyses etc.); however, the most promising route for identification of missing low-frequency and small-effect variants lies through combining biological functional evidence with statistical genetic evidence [33].
Here, we used such “systems genetics” approach, combining gene expression profiling in an animal model of myopia, statistical evidence for association with myopia from two GWAS studies, and functional evidence from a gene-targeted mouse model, to identify APLP2 as one of the “missing” myopia genes. APLP2 was first found to be differentially expressed in the retina of monkeys with experimentally-induced myopia. We then found that numerous SNPs within the 5’-end of APLP2 were associated with refractive error development in children and adults. Furthermore, Aplp2 also strongly influenced refractive eye development and myopia susceptibility in the gene-targeted mouse model of myopia. Interestingly, APLP2 is also localized within a broad suggestive myopia locus with LOD score 3.2 identified by Hammond et al. on chromosome 11q23-24 [93].
APLP2 was first identified as a homologue of the amyloid beta (A4) precursor protein (APP) [50] and was assigned to the human chromosome 11q23-25 [49] and the proximal region of mouse chromosome 9 [47]. It was also found that the expression pattern of APLP2 resembles that of APP in the brain and throughout the body, with particularly high expression in neurons of the central and peripheral nervous system [50,94]. The biological role of APLP2 was investigated using gene-targeted mouse mutants. Aplp2 knockout mice were normal in size, fertile, and appeared healthy, whereas 80% of Aplp2/APP double knockout animals died within 24 h after birth and the remaining 20% exhibited difficulty in righting, ataxia, spinning behavior, and a head tilt [45,52,95]. This was contrasted by no lethality or apparent abnormalities in Aplp1/APP double knockouts, suggesting that the Aplp2 plays a key role in neuronal development and function [52,95–97]. In the brain, APLP2 protein has been localized to the presynaptic active zone of neuronal axons in close proximity to the synaptic vesicles [36]. Consistent with these data, it has been reported that the lack of an Aplp2/APP complex results in reduced expression of vesicular glutamate transporter 2 (VGLUT2) and a defect in synaptic transmission [40,41], as well as reduced spatial learning and synaptic plasticity [37–39]. These effects of Aplp2 on neuronal function have been suggested to be mediated by its role in the subtle modulation of neurotransmitter release [36,39]. Our observation in mice that APLP2 modulates the electrophysiological properties of the retina is consistent with its role in synaptic transmission. We found that lack of Aplp2 led to a significant dose-dependent suppression of the b-wave and oscillatory potentials of the ERG. Considering that oscillatory potentials are primarily generated by the amacrine cells [56,57], which also modulate the amplitude of the b-wave generated by the bipolar cells [58–66], the ERG data suggest that Aplp2 influences refractive eye development by modulating the function of amacrine cells. Interestingly, this is consistent with a previously suggested role for amacrine cells in the regulation of refractive eye development [98–108]. The involvement of Aplp2 in the regulation of retinal processing at the level of amacrine cells is further corroborated by our findings that Aplp2 is expressed in the glycinergic amacrine cells of the retina, which provide feed-forward and feedback inhibition in the retina and play important role in contrast processing [109–115].
Retinal blur associated with inaccurate accommodation during nearwork (so called accommodative lag) and peripheral hyperopic defocus have been hypothesized to be the driving force behind myopia progression in children [116–118]. We found that Aplp2 modulated sensitivity to retinal image degradation induced by form-deprivation in a dose-dependent manner, yet, lack of Aplp2 did not cause an observable change in visual acuity. Thus, our data, in conjunction with the published data on the role of APLP2 in neuronal function, suggest that APLP2 most likely modulates sensitivity to the degradation of retinal images by regulating the processing of contrast by the retina via modulation of synaptic transmission at the level of glycinergic amacrines. The reduced susceptibility to myopia in mice lacking Aplp2 makes lowering the level of APLP2 in the retina via gene therapy an appealing future direction for therapeutic intervention in human myopia.
In summary, we have identified APLP2 as a novel gene involved in refractive eye development and associated with human myopia. The role of APLP2 in human myopia is supported by several lines of evidence, which suggest that genetic variation at the APLP2 promoter region may influence APLP2 expression in the inner retina and, in turn, may modulate synaptic transmission at the level of amacrine cells, leading to alterations in refractive development. Consistent with its important role in neuronal development and function, APLP2 appears to have been subject to intense evolutionary pressure evidenced by 97.7% DNA sequence conservation of the gene between humans and mice. Our findings that naturally occurring genetic variation at the APLP2 locus was associated with myopia only in children who spent an above-average time reading and observations of an analogous gene-environment interaction between Aplp2 and visual input in mice also imply a high level of evolutionary conservation for the pathways underlying refractive eye development. Further work will be required to pinpoint the causal variant(s) at the APLP2 locus that determine susceptibility to myopia, and to elucidate whether (as suggested by the location of the most strongly associated variant in the promoter region of the gene) they alter the level of APLP2 expression. Future functional studies will also need to explore the role of APLP2 in synaptic transmission at the level of amacrine cells and its role in defocus processing.
Materials and Methods
Tests for association with genetic variants at the APLP2 locus in children
To investigate whether naturally-occurring genetic variation at the APLP2 locus influences refractive development in children, data from an existing British birth cohort were examined. The Avon Longitudinal Study of Parents and Children (ALSPAC) recruited 14,541 pregnant women resident in Avon, UK with expected dates of delivery 1st April 1991 to 31st December 1992. Of the initial 14,541 pregnancies, 13,988 children were alive at 1 year. Data collected included self-completion questionnaires sent to the mother, to her partner and after age 5 to the child; direct assessments and interviews in a research clinic; biological samples and linkage to school and hospital records. The original cohort was largely representative of the UK 1991 Census; however, there was trend for greater loss at follow-up for families of low socioeconomic status and of non-White ethnic origin [119]. Ethical approval for the study was obtained from the ALSPAC Law and Ethics committee and the three local research-ethics committees.
Refractive error was assessed using non-cycloplegic autorefraction at research clinics attended when children were approximately 7½, 10½, 11½, 12½ and 15½ years of age [120]. DNA samples from the participants were genotyped on Illumina HumanHap 550 bead arrays [121]. Data were available for a total of 464,311 autosomal SNPs that passed quality control filters [121] in 8,365 individuals of European ancestry (as demonstrated by clustering with HapMap CEU individuals upon multidimensional scaling analysis). SNPs at non-genotyped loci were imputed with MACH,[122] using the 1000-genomes project GIANT consortium November 2010 data release as a reference panel. For attendees at the 15½ year clinic (n = 3,819), association between non-cycloplegic refractive error and imputed genotype dosage was tested using mach2qtl for SNPs within 100 kb of the APLP2 gene. Age and sex were included as covariates in the analysis. Permutation testing was used to generate empirical p-values that accounted for multiple testing and for LD between markers. It was carried out by assigning subjects a new phenotype, sampled randomly without replacement from the true list of phenotypes, and repeating the tests for association between the new trait and the genotype for all SNPs in the region. From 1000 such permutations, the probability of observing a p-value as low as that found in the real dataset was estimated. To test for an excess of low p-values, the probability of observing the 5th percentile p-value from the real dataset was estimated from the 5th percentile p-values observed in the 1000 permutations.
To explore whether the most strongly associated SNP at the APLP2 locus, rs188663068, acted early or late in childhood, the imputed genotype for this SNP was included as a fixed effect term in a linear mixed model of childhood refractive error “trajectory” in ALSPAC participants (note that because of the very low risk allele frequency (RAF) of rs188663068, the single subject who was homozygous for the risk allele (genotype AA) was re-coded as a heterozygote (GA). The model also included sex, age, age2 and age3 as fixed effects, while refractive error over the age range 7½ to 15½ years and a linear age term were modelled as random effects. Data were included for 5,200 ALSPAC participants for whom non-cycloplegic autorefraction readings had been obtained on at least 3 occasions (specifically, there were 833 subjects with data available from 3 visits, 1,696 with data from 4 visits, and 2,671 with data from all 5 visits). Linear mixed modelling was performed using the lme function in R.
More complex refraction trajectory models were constructed by including additional predictor variables. Time spent reading was ascertained from a questionnaire completed by the mother when the ALSPAC participants were aged approximately 8½ years as previously described [120]. In response to the question “On normal days in school holidays, how much time on average does your child spend each day reading books for pleasure”, children were classified as either spending a “high” (response “1–2 hours” or “3 or more hours” per day) or “low” (response “None at all” or “1 hour or less”) amount of time reading. The time reading variable was coded as “low” = 0 (reference) and “high” = 1. There were 2,775 and 1,686 subjects in the “low” and “high” subsets, respectively (note that information on time spent reading was missing for 739 of the 5,200 participants in the refraction trajectory sample). Although time spent reading was sampled at only a single age-point, reading behaviour may track forward as children get older. Therefore, the time spent reading variable’s predictive capacity may stem from capturing inter-subject variation not only at the age of 8–9 years, but also to some extent inter-subject variation in reading behaviour at older ages. Time spent outdoors was gauged from a separate item on the same questionnaire: “On a school weekday, how much time on average does your child spend each day out of doors in summer?”. Children were classified as spending a “high” amount of time outdoors if the response was “1–2 hours” or “3 or more hours”, and as “low” otherwise. Note that this questionnaire response was selected for the present study instead of a closely-related one used previously [120], since the former provided an approximately equal split of the sample, while the latter variable resulted in an ~1 : 9 ratio of subjects classified as spending a low versus high amount of time outdoors, and thus despite its slightly greater predictive discrimination of incident myopia, it would have led to very small numbers of subjects in the “low time outdoors + GA rs188663068 genotype” group. The time outdoors variable was coded as “low” = 0 (reference, n = 2,349) and “high” = 1 (n = 2,145) and there were 706 children with missing information. Sex was not significantly associated with refractive error in the refraction trajectory analyses and so was dropped from the models.
To confirm the refraction trajectory results, association between rs188663068 genotype (coded GG = 0, GA = 1) and refractive error was also analyzed using linear and logistic regression for subjects attending the ALSPAC research clinic; targeting the children when aged 15 years. All children with information available were included in these models to maximise precision of risk estimates. Time spent reading and time spent outdoors were included, separately, as predictors, using the coding scheme described above, in models with and without an interaction term (rs188663068 genotype x time reading, etc.). Sex was not significantly associated with refractive error in these linear regression analyses and so was dropped from the models. Age-at-baseline was not included since, being a birth cohort, the age interval was narrow at each target age. As in the refraction trajectory analyses, the single subject with rs188663068 genotype AA was re-coded as GA.
Tests for association with genetic variants at the APLP2 locus in adults
Genetic variants at the APLP2 locus were examined using meta-analyzed data from the international genome-wide association study (GWAS) of refractive error carried out by the Consortium for Refractive Error and Myopia (CREAM) [30]. The CREAM meta-analysis included data from 32 studies: 1958 British Birth Cohort, ALSPAC (mothers), ANZRAG, AREDS1a1b, AREDS1c, Beijing Eye Study, BMES, CROATIA-Korcula, CROATIA-Split, CROATIA-Vis, DCCT, EGCUT, ERF, FECD, FITSA, Framingham, GHS 1, GHS 2, KORA, OGP Talana, ORCADES, RS1, RS2, RS3, SCES, SIMES, SINDI, SP2, TEST/BATS, TwinsUK, WESDR, and YFS. Each study received prior approval from its local medical ethics committee, and written informed consent was obtained from all participants in accordance with the tenets of the Declaration of Helsinki. The age of subjects in each sample ranged from 31.4 to 79.9 years and, apart from 3 samples, contained an approximately equal split of males/females. Twenty-seven samples comprised of subjects of European ancestry, while 5 were of Asian ancestry. GWAS analyses were carried out for spherical equivalent refractive error (dependent variable) with genotype dosage, age and sex included as independent variables, and meta-analysis was done under a random-effects model, as described [30]. SNPs within 100 kb of the APLP2 gene were evaluated. Permutation-based analysis to correct for multiple testing could not be carried out for the CREAM GWAS dataset since we did not have access to the raw genotypes. Therefore, for SNPs within 100 kb of APLP2, the distribution of CREAM p-values inside versus outside the region showing strong association in the ALSPAC sample (hg19 chr 11 : 129904497–129971498; hg18 chr 11 : 129409707–129476708) was compared using the two-sample Kolmogorov-Smirnov test.
Aplp2 knockout mice
Aplp2 knockout mice (B6.129S7-Aplp2tm1Dbo/J) were obtained from the Jackson Laboratory (Bar Harbor, ME) as heterozygotes and were maintained as an in-house breeding colony on a C57BL/6J background, which was shown not to carry Rd mutations that cause retinal degeneration in mice [123]. To generate homozygous (Aplp2-/-), heterozygous (Aplp2+/-) and wild-type (Aplp2+/+) animals for the experiments, heterozygous males and females were bred and resulting offspring were genotyped as previously described [45] to identify animals of different genotypes. Only littermates were used for all experiments to ensure isogenic genetic background. All animals received water and food ad libitum. All mouse procedures adhered to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research and were approved by the Columbia University Institutional Animal Care and Use Committee (Protocol #AAAK2700). Animals were anesthetized via intraperitoneal injection of pentobarbital (50 mg/kg), or via intraperitoneal injection of ketamine (90 mg/kg) and xylazine (10 mg/kg). Animals were euthanized by cervical dislocation while under full surgical anesthesia.
Analysis of refractive eye development in Aplp2 knockout mice
Visually guided emmetropization normally results in children who are born myopic becoming less myopic and children who are born hyperopic becoming less hyperopic during the early postnatal period [1]. In mice, both the variability and magnitude of the refractive error are reduced during the early postnatal period (P21-P40) indicating emmetropization [124–126]. In C57BL/6J mice, refractive error stabilizes around emmetropia at ~P32. To examine the role of Aplp2 in emmetropization, we analyzed refractive eye development in mice homozygous (Aplp2-/-) and heterozygous (Aplp2+/-) for a null allele of the Aplp2 gene, as well as in the wild-type animals (Aplp2+/+). The refractive state of both left and right eyes was determined on alert animals at P21, P35, and P67 using an automated eccentric infrared photorefractor as previously described [124,127]. The animal to be refracted was immobilized using a restraining platform, and each eye was refracted along the optical axis in dim room light (< 1 lux), 20–30 min. after instilling 1% tropicamide ophthalmic solution (Alcon Laboratories, Inc., Fort Worth, TX) to ensure mydriasis and cycloplegia. Five independent measurement series (~300–600 measurements each) were taken for each eye. The measurements were automatically acquired by the photorefractor every 16 msec. Each successful measurement series (i.e., Purkinje image in the center of the pupil and stable refractive error for at least 5 sec.) was marked by a green LED flash, which was registered by the photorefractor software. Sixty individual measurements from each series, immediately preceding the green LED flash, were combined, and a total of 300 measurements (60 measurements x 5 series = 300 measurements) were used to calculate the refractive error mean and standard deviation.
Analysis of gene-environment interaction between Aplp2 and visual experience in Aplp2 knockout mice
Human population studies revealed that environmental factors, such as nearwork and reading, play important role in the development of myopia [128–131]. These findings were complemented by observations that nearwork and reading are associated with the lag of accommodation, i.e., insufficiently strong accommodative response for near objects, which places the plane of best focus behind the retina (producing slight optical blur) when the subject performs nearwork tasks [116,117]. Optical blur produced by the lag of accommodation is the signal that drives excessive eye growth and causes myopia [116,129,132–135]. Animal studies also demonstrated that excessive eye growth and myopia can be induced in species as diverse as the fish, chicken, tree shrew, monkeys, guinea pig and, most recently, mouse by retinal image degradation or optical blur (recapitulated in animal models by placing a diffuser or a negative lens in front of the eye) [136–143].
To examine the role of Aplp2 in the development of environmentally induced myopia, we analyzed the effect of diffuser-imposed retinal image degradation (visual form deprivation) on refractive eye development in mice homozygous (Aplp2-/-) and heterozygous (Aplp2+/-) for a null allele of the Aplp2 gene, as well as in the wild-type mice (Aplp2+/+). Visual input was degraded in one of the eyes by applying plastic diffusers, and refractive development of the treated eye was compared to that of the contralateral eye, which was not treated with a diffuser, as previously described [126,143]. Diffusers represented low-pass optical filters, which severely degraded the image projected onto the retina by removing high spatial frequency details. Frosted hemispherical plastic diffusers were hand-made using caps from 0.2 ml PCR tubes (Molecular BioProducts, San Diego, CA) and rings made of medical tape (inner diameter 6 mm; outer diameter 8 mm). A cap was frosted with fine sandpaper and attached to a ring with Loctite Super Glue (Henkel Consumer Adhesives, Avon, OH). On the first day of the experiment (P24), animals were anesthetized via intraperitoneal injection of pentobarbital (50 mg/kg), and diffusers were attached to the skin surrounding the right eye with three stitches using size 5–0 ETHILON microsurgical sutures (Ethicon, Somerville, NJ) and reinforced with Vetbond glue (3M Animal Care Products, St. Paul, MN) (the left eye served as a control). Toenails were covered with adhesive tape to prevent mice from removing the diffusers. Animals recovered on a warming pad and were then housed under low-intensity constant light in transparent plastic cages for the duration of the experiment as previously described [126,143]. Following 21 days of visual form deprivation (from P24 through P45), diffusers were removed and refractive status of both treated and control eyes was assessed using an automated eccentric infrared photorefractor as previously described [144]. The interocular difference in refraction between the treated and contralateral control eye served as an indication of the extent of induced myopia.
Analysis of visual acuity and contrast sensitivity in Aplp2 knockout mice
To examine the role of Aplp2 in the overall visual function, we compared visual acuity and contrast-sensitivity in the Aplp2 knockout mice (Aplp2-/-), mice heterozygous (Aplp2+/-) for a null allele of the Aplp2 gene, and in the wild-type littermates (Aplp2+/+). Both visual acuity and contrast sensitivity were measured at P80 using a virtual optomotor system (Mouse OptoMotry System, Cerebral Mechanics, Medicine Hat, AB Canada), as previously described [145]. Briefly, the animal to be tested was placed on a platform surrounded by four computer screens displaying a virtual cylinder comprising a vertical sine wave grating in 3D coordinate space. The OptoMotry software controlled the speed of rotation, direction of rotation, the frequency of the grating and its contrast.
To measure visual acuity, the initial spatial frequency of the grating was set at 0.1 cycles/degree and the contrast was set at maximum. The frequency was then systematically increased using staircase procedure until the maximum spatial frequency capable of eliciting a response (visual acuity) was determined. The staircase procedure was such that 3 correct answers in a row advanced it to a higher spatial frequency, while 1 wrong answer returned it to a lower frequency.
Contrast sensitivity function was measured at seven spatial frequencies, i.e. 0.033, 0.064, 0.083, 0.119, 0.172, 0.244, and 0.347 cycles/degree, using the staircase procedure described above. The contrast sensitivity at each frequency was calculated as a reciprocal of the contrast threshold, which was calculated as a Michelson contrast from the screen luminances ( Imax-IminImax+Imin ; Iwhite = 208.25 cd/m2, Iblack = 0.21 cd/m2).
Analysis of electrophysiological properties of the mouse retina
Dark-adapted electroretinograms (ERGs) are particularly sensitive to changes in the inner retina [56]; therefore, to assess the effect of Aplp2 on the electrophysiological properties of various neuronal populations in the retina, we analyzed scotopic ERGs in the Aplp2 knockout (Aplp2-/-) mice, mice heterozygous for a null allele of the Aplp2 gene (Aplp2+/- mice), and in the wild-type littermates (Aplp2+/+ mice). Animals to be used for ERG were dark-adapted overnight. Prior to ERG recordings, dark-adapted mice were anesthetized via intraperitoneal injection of ketamine (90 mg/kg) and xylazine (10 mg/kg) and placed on a heating pad. The pad was connected to a rectal probe and thermostat via a feedback circuit, which maintained the body temperature at 37°C. Pupil dilation was achieved by instilling one drop of 1% tropicamide ophthalmic solution (Alcon Laboratories, Inc., Fort Worth, TX) in each eye at the time of anesthesia. Silver-embedded thread corneal recording electrodes were positioned across the apex of each cornea anesthetized with Lidocaine and held in place with 2.5% Goniovisc ophthalmic solution (HUB Pharmaceuticals, Rancho Cucamonga, CA) and optically clear mini contact lenses (Ocuscience, Rolla, MO). Stainless steel sub-dermal needle reference electrodes were placed subcutaneously below each eye along the upper jaw, while the ground electrode was inserted into the base of the tail. ERGs were recorded using Ocuscience rodent ERG system (Rolla, MO). To elicit retinal responses, each eye was presented with 5-msec white-light flashes of increasing intensity produced by a mini-Ganzfeld stimulator. Nine intensities ranging from 0.001 cd•s/m2 to 32 cd•s/m2 were used with the interstimulus interval increasing from 18 sec to 120 sec with the increase in the stimulus intensity (12-sec increase for each step). Responses to three flashes of each intensity were recorded and averaged. ERG data were processed and quantified using ERGVIEW software package (Ocuscience, Rolla, MO).
Analysis of Aplp2 expression in the eye
To identify the tissues and cell types in which Aplp2 is expressed, we examined expression of Aplp2 in the eye using in situ hybridization and immunohistochemistry. In situ hybridizations were performed essentially as previously described [146]. Briefly, C57BL/6J mouse eyes were enucleated at P27 and used to prepare eyecups by removing the cornea and lens in ice-cold 1 X PBS. The eyecups were then fixed in 4% paraformaldehyde in 1 X PBS overnight at 4°C, cryoprotected in 30% sucrose in 1 X PBS and embedded in Tissue-Tek O.C.T compound (Sakura Finetek USA, Torrance, CA). 10-μm cryostat sections were incubated with 1 μg/ml Proteinase K in 1 X PBS, washed in 2 mg/ml Glycine in 1 X PBS, incubated with 0.25% acetic anhydride in 0.1 M TEA buffer, and hybridized with digoxigenin(DIG)-labeled cDNA probes followed by incubation with anti-DIG antibodies conjugated with alkaline phosphatase (AP) (Roche Applied Science, Indianapolis, IN). The AP activity was localized and signal was detected using NBT (0.25 mg/ml) and BCIP (0.125 mg/ml) (Roche Applied Science, Indianapolis, IN) as substrates.
For immunohistochemistry, eyecups were prepared as described above, fixed in 2% formaldehyde in 1 X PBS for 4 hours on ice, washed in 1 X PBS, cryoprotected in 30% sucrose in 1 X PBS, and embedded in Tissue-Tek O.C.T compound (Sakura Finetek USA, Torrance, CA). 10-μm cryostat sections were washed with 1 X PBS, blocked with 5% normal goat serum, 5% BSA, 0.1% fish gelatin, 0.1% Triton X-100 and 0.05% Tween 20 in 1 X PBS (blocking buffer) for 1 hour at room temperature, and then incubated with rabbit anti-Aplp2 primary antibodies (D2-II, dilution 1 : 1,000) [46] in blocking buffer overnight at 4°C. The sections were then washed with 0.2% Triton X-100 in 1 X PBS (PBT) and incubated with Alexa-488-conjugated donkey anti-rabbit secondary antibodies (1 : 500, Life Technologies, Grand Island, NY) in blocking buffer for 2 hours at room temperature. After sections were washed in PBT, they were incubated with sheep anti-Chx10 (1 : 200, Abcam, Cambridge, MA), or rabbit anti-Pax6 (1 : 1,000, Abcam, Cambridge, MA), or rabbit anti-GABA (1 : 100, EMD Millipore, Billerica, MA), or rabbit anti-Glycine (1 : 100, EMD Millipore, Billerica, MA) primary antibodies overnight (48 hours for anti-GABA and anti-Glycine antibodies) at 4°C; followed by the washes in PBT and incubation with Alexa-594-conjugated donkey anti-sheep (1 : 500, Life Technologies, Grand Island, NY) or donkey anti-rabbit (1 : 500, Life Technologies, Grand Island, NY) secondary antibodies in blocking buffer for 2 hours at room temperature. The slides were then again washed in PBT, incubated with 300 nM DAPI in 1 X PBS, and mounted in ProLong Gold antifade mountant (Life Technologies, Grand Island, NY). The colocalization between Aplp2 and other antigens was examined and image capture was performed using laser scanning confocal microscope Leica TCS SP5 (Leica Microsystems, Buffalo Grove, IL) and the manufacturer’s software.
Supporting Information
{kind=link}
{kind=link}
Zdroje
1. Oyster CW (1999) The human eye: structure and function. Sunderland, MA: Sinauer Associates.
2. Wallman J, Winawer J (2004) Homeostasis of eye growth and the question of myopia. Neuron 43 : 447–468. 15312645
3. Pararajasegaram R (1999) VISION 2020-the right to sight: from strategies to action. Am J Ophthalmol 128 : 359–360. 10511033
4. Vitale S, Ellwein L, Cotch MF, Ferris FL 3rd, Sperduto R (2008) Prevalence of refractive error in the United States, 1999–2004. Arch Ophthalmol 126 : 1111–1119. doi: 10.1001/archopht.126.8.1111 18695106
5. Lin LL, Shih YF, Hsiao CK, Chen CJ (2004) Prevalence of myopia in Taiwanese schoolchildren: 1983 to 2000. Ann Acad Med Singapore 33 : 27–33.
6. Lam CS, Goldschmidt E, Edwards MH (2004) Prevalence of myopia in local and international schools in Hong Kong. Optom Vis Sci 81 : 317–322. 15181356
7. Pesudovs K, Garamendi E, Elliott DB (2006) A quality of life comparison of people wearing spectacles or contact lenses or having undergone refractive surgery. J Refract Surg 22 : 19–27. 16447932
8. Rose K, Harper R, Tromans C, Waterman C, Goldberg D, et al. (2000) Quality of life in myopia. Br J Ophthalmol 84 : 1031–1034. 10966960
9. Takashima T, Yokoyama T, Futagami S, Ohno-Matsui K, Tanaka H, et al. (2001) The quality of life in patients with pathologic myopia. Jpn J Ophthalmol 45 : 84–92. 11163050
10. Alexander LJ (1994) Primary care of the posterior segment. Connecticut: Appleton & Lange.
11. Saw SM, Gazzard G, Shih-Yen EC, Chua WH (2005) Myopia and associated pathological complications. Ophthalmic Physiol Opt 25 : 381–391. 16101943
12. Flitcroft DI (2012) The complex interactions of retinal, optical and environmental factors in myopia aetiology. Prog Retin Eye Res 31 : 622–660. doi: 10.1016/j.preteyeres.2012.06.004 22772022
13. Pizzarello L, Abiose A, Ffytche T, Duerksen R, Thulasiraj R, et al. (2004) VISION 2020: The Right to Sight: a global initiative to eliminate avoidable blindness. Arch Ophthalmol 122 : 615–620. 15078680
14. Morgan IG (2003) The biological basis of myopic refractive error. Clin Exp Optom 86 : 276–288. 14558849
15. Young TL (2009) Molecular genetics of human myopia: an update. Optom Vis Sci 86: E8–E22. doi: 10.1097/OPX.0b013e3181940655 19104467
16. Baird PN, Schache M, Dirani M (2010) The GEnes in Myopia (GEM) study in understanding the aetiology of refractive errors. Prog Retin Eye Res 29 : 520–542. doi: 10.1016/j.preteyeres.2010.05.004 20576483
17. Wojciechowski R (2011) Nature and nurture: the complex genetics of myopia and refractive error. Clin Genet 79 : 301–320. doi: 10.1111/j.1399-0004.2010.01592.x 21155761
18. Verhoeven VJ, Buitendijk GH, Consortium for Refractive E, Myopia, Rivadeneira F, et al. (2013) Education influences the role of genetics in myopia. Eur J Epidemiol 28 : 973–980. doi: 10.1007/s10654-013-9856-1 24142238
19. Dirani M, Chamberlain M, Shekar SN, Islam AF, Garoufalis P, et al. (2006) Heritability of refractive error and ocular biometrics: the Genes in Myopia (GEM) twin study. Invest Ophthalmol Vis Sci 47 : 4756–4761. 17065484
20. Lopes MC, Andrew T, Carbonaro F, Spector TD, Hammond CJ (2009) Estimating heritability and shared environmental effects for refractive error in twin and family studies. Invest Ophthalmol Vis Sci 50 : 126–131. doi: 10.1167/iovs.08-2385 18757506
21. Peet JA, Cotch MF, Wojciechowski R, Bailey-Wilson JE, Stambolian D (2007) Heritability and familial aggregation of refractive error in the Old Order Amish. Invest Ophthalmol Vis Sci 48 : 4002–4006. 17724179
22. Klein AP, Suktitipat B, Duggal P, Lee KE, Klein R, et al. (2009) Heritability analysis of spherical equivalent, axial length, corneal curvature, and anterior chamber depth in the Beaver Dam Eye Study. Arch Ophthalmol 127 : 649–655. doi: 10.1001/archophthalmol.2009.61 19433716
23. Chen CY, Scurrah KJ, Stankovich J, Garoufalis P, Dirani M, et al. (2007) Heritability and shared environment estimates for myopia and associated ocular biometric traits: the Genes in Myopia (GEM) family study. Hum Genet 121 : 511–520. 17205325
24. Wojciechowski R, Congdon N, Bowie H, Munoz B, Gilbert D, et al. (2005) Heritability of refractive error and familial aggregation of myopia in an elderly American population. Invest Ophthalmol Vis Sci 46 : 1588–1592. 15851555
25. Solouki AM, Verhoeven VJ, van Duijn CM, Verkerk AJ, Ikram MK, et al. (2010) A genome-wide association study identifies a susceptibility locus for refractive errors and myopia at 15q14. Nat Genet 42 : 897–901. doi: 10.1038/ng.663 20835239
26. Hysi PG, Young TL, Mackey DA, Andrew T, Fernandez-Medarde A, et al. (2010) A genome-wide association study for myopia and refractive error identifies a susceptibility locus at 15q25. Nat Genet 42 : 902–905. doi: 10.1038/ng.664 20835236
27. Li Z, Qu J, Xu X, Zhou X, Zou H, et al. (2011) A genome-wide association study reveals association between common variants in an intergenic region of 4q25 and high-grade myopia in the Chinese Han population. Hum Mol Genet 20 : 2861–2868. doi: 10.1093/hmg/ddr169 21505071
28. Shi Y, Qu J, Zhang D, Zhao P, Zhang Q, et al. (2011) Genetic variants at 13q12.12 are associated with high myopia in the han chinese population. Am J Hum Genet 88 : 805–813. doi: 10.1016/j.ajhg.2011.04.022 21640322
29. Li YJ, Goh L, Khor CC, Fan Q, Yu M, et al. (2011) Genome-wide association studies reveal genetic variants in CTNND2 for high myopia in Singapore Chinese. Ophthalmology 118 : 368–375. doi: 10.1016/j.ophtha.2010.06.016 21095009
30. Verhoeven VJ, Hysi PG, Wojciechowski R, Fan Q, Guggenheim JA, et al. (2013) Genome-wide meta-analyses of multiancestry cohorts identify multiple new susceptibility loci for refractive error and myopia. Nat Genet 45 : 314–318. doi: 10.1038/ng.2554 23396134
31. Farbrother JE, Kirov G, Owen MJ, Pong-Wong R, Haley CS, et al. (2004) Linkage analysis of the genetic loci for high myopia on 18p, 12q, and 17q in 51 U.K. families. Invest Ophthalmol Vis Sci 45 : 2879–2885. 15326098
32. Gusev A, Bhatia G, Zaitlen N, Vilhjalmsson BJ, Diogo D, et al. (2013) Quantifying missing heritability at known GWAS loci. PLoS Genet 9: e1003993. doi: 10.1371/journal.pgen.1003993 24385918
33. Manolio TA, Collins FS, Cox NJ, Goldstein DB, Hindorff LA, et al. (2009) Finding the missing heritability of complex diseases. Nature 461 : 747–753. doi: 10.1038/nature08494 19812666
34. Eyre-Walker A (2010) Evolution in health and medicine Sackler colloquium: Genetic architecture of a complex trait and its implications for fitness and genome-wide association studies. Proc Natl Acad Sci U S A 107 Suppl 1 : 1752–1756. doi: 10.1073/pnas.0906182107 20133822
35. Park JH, Gail MH, Weinberg CR, Carroll RJ, Chung CC, et al. (2011) Distribution of allele frequencies and effect sizes and their interrelationships for common genetic susceptibility variants. Proc Natl Acad Sci U S A 108 : 18026–18031. doi: 10.1073/pnas.1114759108 22003128
36. Lassek M, Weingarten J, Einsfelder U, Brendel P, Muller U, et al. (2013) Amyloid precursor proteins are constituents of the presynaptic active zone. J Neurochem 127 : 48–56. doi: 10.1111/jnc.12358 23815291
37. Zhang X, Herrmann U, Weyer SW, Both M, Muller UC, et al. (2013) Hippocampal network oscillations in APP/APLP2-deficient mice. PLoS One 8: e61198. doi: 10.1371/journal.pone.0061198 23585881
38. Korte M, Herrmann U, Zhang X, Draguhn A (2012) The role of APP and APLP for synaptic transmission, plasticity, and network function: lessons from genetic mouse models. Exp Brain Res 217 : 435–440. doi: 10.1007/s00221-011-2894-6 22006270
39. Weyer SW, Klevanski M, Delekate A, Voikar V, Aydin D, et al. (2011) APP and APLP2 are essential at PNS and CNS synapses for transmission, spatial learning and LTP. EMBO J 30 : 2266–2280. doi: 10.1038/emboj.2011.119 21522131
40. Schrenk-Siemens K, Perez-Alcala S, Richter J, Lacroix E, Rahuel J, et al. (2008) Embryonic stem cell-derived neurons as a cellular system to study gene function: lack of amyloid precursor proteins APP and APLP2 leads to defective synaptic transmission. Stem Cells 26 : 2153–2163. doi: 10.1634/stemcells.2008-0010 18535156
41. Herard AS, Besret L, Dubois A, Dauguet J, Delzescaux T, et al. (2006) siRNA targeted against amyloid precursor protein impairs synaptic activity in vivo. Neurobiol Aging 27 : 1740–1750. 16337035
42. Yang G, Gong YD, Gong K, Jiang WL, Kwon E, et al. (2005) Reduced synaptic vesicle density and active zone size in mice lacking amyloid precursor protein (APP) and APP-like protein 2. Neurosci Lett 384 : 66–71. 15919150
43. Wang P, Yang G, Mosier DR, Chang P, Zaidi T, et al. (2005) Defective neuromuscular synapses in mice lacking amyloid precursor protein (APP) and APP-Like protein 2. J Neurosci 25 : 1219–1225. 15689559
44. Leach R, Ko M, Krawetz SA (1999) Assignment of amyloid-precursor-like protein 2 gene (APLP2) to 11q24 by fluorescent in situ hybridization. Cytogenet Cell Genet 87 : 215–216. 10702673
45. von Koch CS, Zheng H, Chen H, Trumbauer M, Thinakaran G, et al. (1997) Generation of APLP2 KO mice and early postnatal lethality in APLP2/APP double KO mice. Neurobiol Aging 18 : 661–669. 9461064
46. Thinakaran G, Kitt CA, Roskams AJ, Slunt HH, Masliah E, et al. (1995) Distribution of an APP homolog, APLP2, in the mouse olfactory system: a potential role for APLP2 in axogenesis. J Neurosci 15 : 6314–6326. 7472397
47. von Koch CS, Lahiri DK, Mammen AL, Copeland NG, Gilbert DJ, et al. (1995) The mouse APLP2 gene. Chromosomal localization and promoter characterization. J Biol Chem 270 : 25475–25480. 7592716
48. Sandbrink R, Masters CL, Beyreuther K (1994) Complete nucleotide and deduced amino acid sequence of rat amyloid protein precursor-like protein 2 (APLP2/APPH): two amino acids length difference to human and murine homologues. Biochim Biophys Acta 1219 : 167–170. 8086458
49. von der Kammer H, Loffler C, Hanes J, Klaudiny J, Scheit KH, et al. (1994) The gene for the amyloid precursor-like protein APLP2 is assigned to human chromosome 11q23-q25. Genomics 20 : 308–311. 8020984
50. Wasco W, Gurubhagavatula S, Paradis MD, Romano DM, Sisodia SS, et al. (1993) Isolation and characterization of APLP2 encoding a homologue of the Alzheimer's associated amyloid beta protein precursor. Nat Genet 5 : 95–100. 8220435
51. Cousins SL, Dai W, Stephenson FA (2015) APLP1 and APLP2, members of the APP family of proteins, behave similarly to APP in that they associate with NMDA receptors and enhance NMDA receptor surface expression. J Neurochem.
52. Aydin D, Weyer SW, Muller UC (2012) Functions of the APP gene family in the nervous system: insights from mouse models. Exp Brain Res 217 : 423–434. doi: 10.1007/s00221-011-2861-2 21931985
53. Tkatchenko AV, Walsh PA, Tkatchenko TV, Gustincich S, Raviola E (2006) Form deprivation modulates retinal neurogenesis in primate experimental myopia. Proc Natl Acad Sci U S A 103 : 4681–4686. 16537371
54. Subramanian A, Kuehn H, Gould J, Tamayo P, Mesirov JP (2007) GSEA-P: a desktop application for Gene Set Enrichment Analysis. Bioinformatics 23 : 3251–3253. 17644558
55. Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, et al. (2005) Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A 102 : 15545–15550. 16199517
56. Wachtmeister L (1998) Oscillatory potentials in the retina: what do they reveal. Prog Retin Eye Res 17 : 485–521. 9777648
57. Dong CJ, Agey P, Hare WA (2004) Origins of the electroretinogram oscillatory potentials in the rabbit retina. Vis Neurosci 21 : 533–543. 15579219
58. Frumkes TE, Nelson R, Pflug R (1995) Functional role of GABA in cat retina: II. Effects of GABAA antagonists. Vis Neurosci 12 : 651–661. 8527367
59. Green DG, Kapousta-Bruneau NV (1999) A dissection of the electroretinogram from the isolated rat retina with microelectrodes and drugs. Vis Neurosci 16 : 727–741. 10431921
60. Hood DC, Birch DG (1996) Beta wave of the scotopic (rod) electroretinogram as a measure of the activity of human on-bipolar cells. J Opt Soc Am A Opt Image Sci Vis 13 : 623–633. 8627419
61. Sharma S, Ball SL, Peachey NS (2005) Pharmacological studies of the mouse cone electroretinogram. Vis Neurosci 22 : 631–636. 16332274
62. Sieving PA, Murayama K, Naarendorp F (1994) Push-pull model of the primate photopic electroretinogram: a role for hyperpolarizing neurons in shaping the b-wave. Vis Neurosci 11 : 519–532. 8038126
63. Kapousta-Bruneau NV (2000) Opposite effects of GABA(A) and GABA(C) receptor antagonists on the b-wave of ERG recorded from the isolated rat retina. Vision Res 40 : 1653–1665. 10814754
64. Dong CJ, Hare WA (2002) GABAc feedback pathway modulates the amplitude and kinetics of ERG b-wave in a mammalian retina in vivo. Vision Res 42 : 1081–1087. 11997047
65. Dong CJ, Hare WA (2000) Contribution to the kinetics and amplitude of the electroretinogram b-wave by third-order retinal neurons in the rabbit retina. Vision Res 40 : 579–589. 10824262
66. Lewis A, Wilson N, Stearns G, Johnson N, Nelson R, et al. (2011) Celsr3 is required for normal development of GABA circuits in the inner retina. PLoS Genet 7: e1002239. doi: 10.1371/journal.pgen.1002239 21852962
67. Young TL, Ronan SM, Drahozal LA, Wildenberg SC, Alvear AB, et al. (1998) Evidence that a locus for familial high myopia maps to chromosome 18p. Am J Hum Genet 63 : 109–119. 9634508
68. Young TL, Ronan SM, Alvear AB, Wildenberg SC, Oetting WS, et al. (1998) A second locus for familial high myopia maps to chromosome 12q. Am J Hum Genet 63 : 1419–1424. 9792869
69. Naiglin L, Gazagne C, Dallongeville F, Thalamas C, Idder A, et al. (2002) A genome wide scan for familial high myopia suggests a novel locus on chromosome 7q36. J Med Genet 39 : 118–124. 11836361
70. Lam DS, Tam PO, Fan DS, Baum L, Leung YF, et al. (2003) Familial high myopia linkage to chromosome 18p. Ophthalmologica 217 : 115–118. 12592049
71. Paluru P, Ronan SM, Heon E, Devoto M, Wildenberg SC, et al. (2003) New locus for autosomal dominant high myopia maps to the long arm of chromosome 17. Invest Ophthalmol Vis Sci 44 : 1830–1836. 12714612
72. Stambolian D, Ibay G, Reider L, Dana D, Moy C, et al. (2004) Genomewide linkage scan for myopia susceptibility loci among Ashkenazi Jewish families shows evidence of linkage on chromosome 22q12. Am J Hum Genet 75 : 448–459. 15273935
73. Zhang Q, Guo X, Xiao X, Jia X, Li S, et al. (2005) A new locus for autosomal dominant high myopia maps to 4q22-q27 between D4S1578 and D4S1612. Mol Vis 11 : 554–560. 16052171
74. Paluru PC, Nallasamy S, Devoto M, Rappaport EF, Young TL (2005) Identification of a novel locus on 2q for autosomal dominant high-grade myopia. Invest Ophthalmol Vis Sci 46 : 2300–2307. 15980214
75. Stambolian D, Ciner EB, Reider LC, Moy C, Dana D, et al. (2005) Genome-wide scan for myopia in the Old Order Amish. Am J Ophthalmol 140 : 469–476. 16084785
76. Zhang Q, Guo X, Xiao X, Jia X, Li S, et al. (2006) Novel locus for X linked recessive high myopia maps to Xq23-q25 but outside MYP1. J Med Genet 43: e20. 16648373
77. Nallasamy S, Paluru PC, Devoto M, Wasserman NF, Zhou J, et al. (2007) Genetic linkage study of high-grade myopia in a Hutterite population from South Dakota. Mol Vis 13 : 229–236. 17327828
78. Yu ZQ, Li YB, Huang CX, Chu RY, Hu DN, et al. (2007) [A genome-wide screening for pathological myopia suggests a novel locus on chromosome 15q12–13]. Zhonghua Yan Ke Za Zhi 43 : 233–238. 17605906
79. Paget S, Julia S, Vitezica ZG, Soler V, Malecaze F, et al. (2008) Linkage analysis of high myopia susceptibility locus in 26 families. Mol Vis 14 : 2566–2574. 19122830
80. Ciner E, Wojciechowski R, Ibay G, Bailey-Wilson JE, Stambolian D (2008) Genomewide scan of ocular refraction in African-American families shows significant linkage to chromosome 7p15. Genet Epidemiol 32 : 454–463. doi: 10.1002/gepi.20318 18293391
81. Lam CY, Tam PO, Fan DS, Fan BJ, Wang DY, et al. (2008) A genome-wide scan maps a novel high myopia locus to 5p15. Invest Ophthalmol Vis Sci 49 : 3768–3778. doi: 10.1167/iovs.07-1126 18421076
82. Schache M, Chen CY, Pertile KK, Richardson AJ, Dirani M, et al. (2009) Fine mapping linkage analysis identifies a novel susceptibility locus for myopia on chromosome 2q37 adjacent to but not overlapping MYP12. Mol Vis 15 : 722–730. 19365569
83. Yang Z, Xiao X, Li S, Zhang Q (2009) Clinical and linkage study on a consanguineous Chinese family with autosomal recessive high myopia. Mol Vis 15 : 312–318. 19204786
84. Nishizaki R, Ota M, Inoko H, Meguro A, Shiota T, et al. (2009) New susceptibility locus for high myopia is linked to the uromodulin-like 1 (UMODL1) gene region on chromosome 21q22.3. Eye (Lond) 23 : 222–229.
85. Ciner E, Ibay G, Wojciechowski R, Dana D, Holmes TN, et al. (2009) Genome-wide scan of African-American and white families for linkage to myopia. Am J Ophthalmol 147 : 512–517 e512. doi: 10.1016/j.ajo.2008.09.004 19026404
86. Li YJ, Guggenheim JA, Bulusu A, Metlapally R, Abbott D, et al. (2009) An international collaborative family-based whole-genome linkage scan for high-grade myopia. Invest Ophthalmol Vis Sci 50 : 3116–3127. doi: 10.1167/iovs.08-2781 19324860
87. Nakanishi H, Yamada R, Gotoh N, Hayashi H, Yamashiro K, et al. (2009) A genome-wide association analysis identified a novel susceptible locus for pathological myopia at 11q24.1. PLoS Genet 5: e1000660. doi: 10.1371/journal.pgen.1000660 19779542
88. Ma JH, Shen SH, Zhang GW, Zhao DS, Xu C, et al. (2010) Identification of a locus for autosomal dominant high myopia on chromosome 5p13.3-p15.1 in a Chinese family. Mol Vis 16 : 2043–2054. 21042559
89. Kiefer AK, Tung JY, Do CB, Hinds DA, Mountain JL, et al. (2013) Genome-wide analysis points to roles for extracellular matrix remodeling, the visual cycle, and neuronal development in myopia. PLoS Genet 9: e1003299. doi: 10.1371/journal.pgen.1003299 23468642
90. Lango Allen H, Estrada K, Lettre G, Berndt SI, Weedon MN, et al. (2010) Hundreds of variants clustered in genomic loci and biological pathways affect human height. Nature 467 : 832–838. doi: 10.1038/nature09410 20881960
91. Hammond CJ, Snieder H, Gilbert CE, Spector TD (2001) Genes and environment in refractive error: the twin eye study. Invest Ophthalmol Vis Sci 42 : 1232–1236. 11328732
92. Lyhne N, Sjolie AK, Kyvik KO, Green A (2001) The importance of genes and environment for ocular refraction and its determiners: a population based study among 20–45 year old twins. Br J Ophthalmol 85 : 1470–1476. 11734523
93. Hammond CJ, Andrew T, Mak YT, Spector TD (2004) A susceptibility locus for myopia in the normal population is linked to the PAX6 gene region on chromosome 11: a genomewide scan of dizygotic twins. Am J Hum Genet 75 : 294–304. 15307048
94. Slunt HH, Thinakaran G, Von Koch C, Lo AC, Tanzi RE, et al. (1994) Expression of a ubiquitous, cross-reactive homologue of the mouse beta-amyloid precursor protein (APP). J Biol Chem 269 : 2637–2644. 8300594
95. Heber S, Herms J, Gajic V, Hainfellner J, Aguzzi A, et al. (2000) Mice with combined gene knock-outs reveal essential and partially redundant functions of amyloid precursor protein family members. J Neurosci 20 : 7951–7963. 11050115
96. Soba P, Eggert S, Wagner K, Zentgraf H, Siehl K, et al. (2005) Homo - and heterodimerization of APP family members promotes intercellular adhesion. EMBO J 24 : 3624–3634. 16193067
97. Herms J, Anliker B, Heber S, Ring S, Fuhrmann M, et al. (2004) Cortical dysplasia resembling human type 2 lissencephaly in mice lacking all three APP family members. EMBO J 23 : 4106–4115. 15385965
98. Stone RA, Laties AM, Raviola E, Wiesel TN (1988) Increase in retinal vasoactive intestinal polypeptide after eyelid fusion in primates. Proc Natl Acad Sci U S A 85 : 257–260. 2448769
99. Seltner RL, Stell WK (1995) The effect of vasoactive intestinal peptide on development of form deprivation myopia in the chick: a pharmacological and immunocytochemical study. Vision Res 35 : 1265–1270. 7610586
100. Fischer AJ, Seltner RL, Stell WK (1997) N-methyl-D-aspartate-induced excitotoxicity causes myopia in hatched chicks. Can J Ophthalmol 32 : 373–377. 9363340
101. Fischer AJ, McGuire JJ, Schaeffel F, Stell WK (1999) Light - and focus-dependent expression of the transcription factor ZENK in the chick retina. Nat Neurosci 2 : 706–712. 10412059
102. Feldkaemper MP, Schaeffel F (2002) Evidence for a potential role of glucagon during eye growth regulation in chicks. Vis Neurosci 19 : 755–766. 12688670
103. Zhong X, Ge J, Smith EL 3rd, Stell WK (2004) Image defocus modulates activity of bipolar and amacrine cells in macaque retina. Invest Ophthalmol Vis Sci 45 : 2065–2074. 15223778
104. Vessey KA, Lencses KA, Rushforth DA, Hruby VJ, Stell WK (2005) Glucagon receptor agonists and antagonists affect the growth of the chick eye: a role for glucagonergic regulation of emmetropization? Invest Ophthalmol Vis Sci 46 : 3922–3931. 16249465
105. Chen JC, Brown B, Schmid KL (2006) Evaluation of inner retinal function in myopia using oscillatory potentials of the multifocal electroretinogram. Vision Res 46 : 4096–4103. 17010409
106. Mathis U, Schaeffel F (2007) Glucagon-related peptides in the mouse retina and the effects of deprivation of form vision. Graefes Arch Clin Exp Ophthalmol 245 : 267–275. 16741711
107. Feldkaemper MP, Neacsu I, Schaeffel F (2009) Insulin acts as a powerful stimulator of axial myopia in chicks. Invest Ophthalmol Vis Sci 50 : 13–23. doi: 10.1167/iovs.08-1702 18599564
108. Ashby R, Kozulin P, Megaw PL, Morgan IG (2010) Alterations in ZENK and glucagon RNA transcript expression during increased ocular growth in chickens. Mol Vis 16 : 639–649. 20405027
109. Wassle H, Heinze L, Ivanova E, Majumdar S, Weiss J, et al. (2009) Glycinergic transmission in the Mammalian retina. Front Mol Neurosci 2 : 6. doi: 10.3389/neuro.02.006.2009 19924257
110. Werblin FS (2010) Six different roles for crossover inhibition in the retina: correcting the nonlinearities of synaptic transmission. Vis Neurosci 27 : 1–8. doi: 10.1017/S0952523810000076 20392301
111. Russell TL, Werblin FS (2010) Retinal synaptic pathways underlying the response of the rabbit local edge detector. J Neurophysiol 103 : 2757–2769. doi: 10.1152/jn.00987.2009 20457864
112. Zhang C, McCall MA (2012) Receptor targets of amacrine cells. Vis Neurosci 29 : 11–29. doi: 10.1017/S0952523812000028 22310370
113. Buldyrev I, Taylor WR (2013) Inhibitory mechanisms that generate centre and surround properties in ON and OFF brisk-sustained ganglion cells in the rabbit retina. J Physiol 591 : 303–325. doi: 10.1113/jphysiol.2012.243113 23045347
114. Buldyrev I, Puthussery T, Taylor WR (2012) Synaptic pathways that shape the excitatory drive in an OFF retinal ganglion cell. J Neurophysiol 107 : 1795–1807. doi: 10.1152/jn.00924.2011 22205648
115. Venkataramani S, Van Wyk M, Buldyrev I, Sivyer B, Vaney DI, et al. (2014) Distinct roles for inhibition in spatial and temporal tuning of local edge detectors in the rabbit retina. PLoS One 9: e88560. doi: 10.1371/journal.pone.0088560 24586343
116. Gwiazda J, Thorn F, Bauer J, Held R (1993) Myopic children show insufficient accommodative response to blur. Invest Ophthalmol Vis Sci 34 : 690–694. 8449687
117. Seidemann A, Schaeffel F (2003) An evaluation of the lag of accommodation using photorefraction. Vision Res 43 : 419–430. 12535999
118. Smith EL 3rd, Hung LF, Huang J (2009) Relative peripheral hyperopic defocus alters central refractive development in infant monkeys. Vision Res 49 : 2386–2392. doi: 10.1016/j.visres.2009.07.011 19632261
119. Boyd A, Golding J, Macleod J, Lawlor DA, Fraser A, et al. (2013) Cohort Profile: the 'children of the 90s'—the index offspring of the Avon Longitudinal Study of Parents and Children. Int J Epidemiol 42 : 111–127. doi: 10.1093/ije/dys064 22507743
120. Guggenheim JA, Northstone K, McMahon G, Ness AR, Deere K, et al. (2012) Time outdoors and physical activity as predictors of incident myopia in childhood: a prospective cohort study. Invest Ophthalmol Vis Sci 53 : 2856–2865. doi: 10.1167/iovs.11-9091 22491403
121. Guggenheim JA, Zhou X, Evans DM, Timpson NJ, McMahon G, et al. (2013) Coordinated genetic scaling of the human eye: shared determination of axial eye length and corneal curvature. Invest Ophthalmol Vis Sci 54 : 1715–1721. doi: 10.1167/iovs.12-10560 23385790
122. Li Y, Willer CJ, Ding J, Scheet P, Abecasis GR (2010) MaCH: using sequence and genotype data to estimate haplotypes and unobserved genotypes. Genet Epidemiol 34 : 816–834. doi: 10.1002/gepi.20533 21058334
123. Mattapallil MJ, Wawrousek EF, Chan CC, Zhao H, Roychoudhury J, et al. (2012) The Rd8 Mutation of the Crb1 Gene Is Present in Vendor Lines of C57BL/6N Mice and Embryonic Stem Cells, and Confounds Ocular Induced Mutant Phenotypes. Invest Ophthalmol Vis Sci 53 : 2921–2927. doi: 10.1167/iovs.12-9662 22447858
124. Tkatchenko TV, Shen Y, Tkatchenko AV (2010) Analysis of postnatal eye development in the mouse with high-resolution small animal magnetic resonance imaging. Invest Ophthalmol Vis Sci 51 : 21–27. doi: 10.1167/iovs.08-2767 19661239
125. Tkatchenko AV, Shen Y, Tkatchenko TV (2012) Genetic background modulates refractive eye development and susceptibility to myopia in the mouse. Invest Ophthalmol Vis Sci 53: E-Abstract 3465.
126. Tkatchenko TV, Shen Y, Braun RD, Bawa G, Kumar P, et al. (2013) Photopic visual input is necessary for emmetropization in mice. Exp Eye Res 115C: 87–95.
127. Tkatchenko TV, Tkatchenko AV (2010) Ketamine-xylazine anesthesia causes hyperopic refractive shift in mice. J Neurosci Methods 193 : 67–71. doi: 10.1016/j.jneumeth.2010.07.036 20813132
128. Parssinen O, Lyyra AL (1993) Myopia and myopic progression among schoolchildren: a three-year follow-up study. Invest Ophthalmol Vis Sci 34 : 2794–2802. 8344801
129. Goss DA (2000) Nearwork and myopia. Lancet 356 : 1456–1457. 11081523
130. Hepsen IF, Evereklioglu C, Bayramlar H (2001) The effect of reading and near-work on the development of myopia in emmetropic boys: a prospective, controlled, three-year follow-up study. Vision Res 41 : 2511–2520. 11483181
131. Saw SM, Chua WH, Hong CY, Wu HM, Chan WY, et al. (2002) Nearwork in early-onset myopia. Invest Ophthalmol Vis Sci 43 : 332–339. 11818374
132. Gwiazda J, Bauer J, Thorn F, Held R (1995) A dynamic relationship between myopia and blur-driven accommodation in school-aged children. Vision Res 35 : 1299–1304. 7610590
133. Abbott ML, Schmid KL, Strang NC (1998) Differences in the accommodation stimulus response curves of adult myopes and emmetropes. Ophthalmic Physiol Opt 18 : 13–20. 9666906
134. Charman WN (1999) Near vision, lags of accommodation and myopia. Ophthalmic Physiol Opt 19 : 126–133. 10615448
135. Gwiazda JE, Hyman L, Norton TT, Hussein ME, Marsh-Tootle W, et al. (2004) Accommodation and related risk factors associated with myopia progression and their interaction with treatment in COMET children. Invest Ophthalmol Vis Sci 45 : 2143–2151. 15223788
136. Shen W, Vijayan M, Sivak JG (2005) Inducing form-deprivation myopia in fish. Invest Ophthalmol Vis Sci 46 : 1797–1803. 15851585
137. Wallman J, Turkel J, Trachtman J (1978) Extreme myopia produced by modest change in early visual experience. Science 201 : 1249–1251. 694514
138. Sherman SM, Norton TT, Casagrande VA (1977) Myopia in the lid-sutured tree shrew (Tupaia glis). Brain Res 124 : 154–157. 843938
139. Wiesel TN, Raviola E (1977) Myopia and eye enlargement after neonatal lid fusion in monkeys. Nature 266 : 66–68. 402582
140. Smith EL 3rd, Hung LF (1999) The role of optical defocus in regulating refractive development in infant monkeys. Vision Res 39 : 1415–1435. 10343811
141. Howlett MH, McFadden SA (2009) Spectacle lens compensation in the pigmented guinea pig. Vision Res 49 : 219–227. doi: 10.1016/j.visres.2008.10.008 18992765
142. Howlett MH, McFadden SA (2006) Form-deprivation myopia in the guinea pig (Cavia porcellus). Vision Res 46 : 267–283. 16139323
143. Tkatchenko TV, Shen Y, Tkatchenko AV (2010) Mouse experimental myopia has features of primate myopia. Invest Ophthalmol Vis Sci 51 : 1297–1303. doi: 10.1167/iovs.09-4153 19875658
144. Schaeffel F, Burkhardt E, Howland HC, Williams RW (2004) Measurement of refractive state and deprivation myopia in two strains of mice. Optom Vis Sci 81 : 99–110. 15127929
145. Prusky GT, Alam NM, Beekman S, Douglas RM (2004) Rapid quantification of adult and developing mouse spatial vision using a virtual optomotor system. Invest Ophthalmol Vis Sci 45 : 4611–4616. 15557474
146. Yang H, Wanner IB, Roper SD, Chaudhari N (1999) An optimized method for in situ hybridization with signal amplification that allows the detection of rare mRNAs. J Histochem Cytochem 47 : 431–446. 10082745
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2015 Číslo 8
Nejčtenější v tomto čísle
- Exon 7 Contributes to the Stable Localization of Xist RNA on the Inactive X-Chromosome
- YAP1 Exerts Its Transcriptional Control via TEAD-Mediated Activation of Enhancers
- SmD1 Modulates the miRNA Pathway Independently of Its Pre-mRNA Splicing Function
- TSPO, a Mitochondrial Outer Membrane Protein, Controls Ethanol-Related Behaviors in