
Diagnosing Severe Falciparum Malaria in Parasitaemic African Children: A Prospective Evaluation of Plasma HRP2 Measurement
Background:
In African children, distinguishing severe falciparum malaria from other severe febrile illnesses with coincidental Plasmodium falciparum parasitaemia is a major challenge. P. falciparum histidine-rich protein 2 (PfHRP2) is released by mature sequestered parasites and can be used to estimate the total parasite burden. We investigated the prognostic significance of plasma PfHRP2 and used it to estimate the malaria-attributable fraction in African children diagnosed with severe malaria.
Methods and Findings:
Admission plasma PfHRP2 was measured prospectively in African children (from Mozambique, The Gambia, Kenya, Tanzania, Uganda, Rwanda, and the Democratic Republic of the Congo) aged 1 month to 15 years with severe febrile illness and a positive P. falciparum lactate dehydrogenase (pLDH)-based rapid test in a clinical trial comparing parenteral artesunate versus quinine (the AQUAMAT trial, ISRCTN 50258054). In 3,826 severely ill children, Plasmadium falciparum PfHRP2 was higher in patients with coma (p = 0.0209), acidosis (p<0.0001), and severe anaemia (p<0.0001). Admission geometric mean (95%CI) plasma PfHRP2 was 1,611 (1,350–1,922) ng/mL in fatal cases (n = 381) versus 1,046 (991–1,104) ng/mL in survivors (n = 3,445, p<0.0001), without differences in parasitaemia as assessed by microscopy. There was a U-shaped association between log10 plasma PfHRP2 and risk of death. Mortality increased 20% per log10 increase in PfHRP2 above 174 ng/mL (adjusted odds ratio [AOR] 1.21, 95%CI 1.05–1.39, p = 0.009). A mechanistic model assuming a PfHRP2-independent risk of death in non-malaria illness closely fitted the observed data and showed malaria-attributable mortality less than 50% with plasma PfHRP2≤174 ng/mL. The odds ratio (OR) for death in artesunate versus quinine-treated patients was 0.61 (95%CI 0.44–0.83, p = 0.0018) in the highest PfHRP2 tertile, whereas there was no difference in the lowest tertile (OR 1.05; 95%CI 0.69–1.61; p = 0.82). A limitation of the study is that some conclusions are drawn from a mechanistic model, which is inherently dependent on certain assumptions. However, a sensitivity analysis of the model indicated that the results were robust to a plausible range of parameter estimates. Further studies are needed to validate our findings.
Conclusions:
Plasma PfHRP2 has prognostic significance in African children with severe falciparum malaria and provides a tool to stratify the risk of “true” severe malaria-attributable disease as opposed to other severe illnesses in parasitaemic African children.
Please see later in the article for the Editors' Summary.
Published in the journal:
. PLoS Med 9(8): e32767. doi:10.1371/journal.pmed.1001297
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pmed.1001297
Summary
Background:
In African children, distinguishing severe falciparum malaria from other severe febrile illnesses with coincidental Plasmodium falciparum parasitaemia is a major challenge. P. falciparum histidine-rich protein 2 (PfHRP2) is released by mature sequestered parasites and can be used to estimate the total parasite burden. We investigated the prognostic significance of plasma PfHRP2 and used it to estimate the malaria-attributable fraction in African children diagnosed with severe malaria.
Methods and Findings:
Admission plasma PfHRP2 was measured prospectively in African children (from Mozambique, The Gambia, Kenya, Tanzania, Uganda, Rwanda, and the Democratic Republic of the Congo) aged 1 month to 15 years with severe febrile illness and a positive P. falciparum lactate dehydrogenase (pLDH)-based rapid test in a clinical trial comparing parenteral artesunate versus quinine (the AQUAMAT trial, ISRCTN 50258054). In 3,826 severely ill children, Plasmadium falciparum PfHRP2 was higher in patients with coma (p = 0.0209), acidosis (p<0.0001), and severe anaemia (p<0.0001). Admission geometric mean (95%CI) plasma PfHRP2 was 1,611 (1,350–1,922) ng/mL in fatal cases (n = 381) versus 1,046 (991–1,104) ng/mL in survivors (n = 3,445, p<0.0001), without differences in parasitaemia as assessed by microscopy. There was a U-shaped association between log10 plasma PfHRP2 and risk of death. Mortality increased 20% per log10 increase in PfHRP2 above 174 ng/mL (adjusted odds ratio [AOR] 1.21, 95%CI 1.05–1.39, p = 0.009). A mechanistic model assuming a PfHRP2-independent risk of death in non-malaria illness closely fitted the observed data and showed malaria-attributable mortality less than 50% with plasma PfHRP2≤174 ng/mL. The odds ratio (OR) for death in artesunate versus quinine-treated patients was 0.61 (95%CI 0.44–0.83, p = 0.0018) in the highest PfHRP2 tertile, whereas there was no difference in the lowest tertile (OR 1.05; 95%CI 0.69–1.61; p = 0.82). A limitation of the study is that some conclusions are drawn from a mechanistic model, which is inherently dependent on certain assumptions. However, a sensitivity analysis of the model indicated that the results were robust to a plausible range of parameter estimates. Further studies are needed to validate our findings.
Conclusions:
Plasma PfHRP2 has prognostic significance in African children with severe falciparum malaria and provides a tool to stratify the risk of “true” severe malaria-attributable disease as opposed to other severe illnesses in parasitaemic African children.
Please see later in the article for the Editors' Summary.
Introduction
Severe falciparum malaria in children presents a major diagnostic challenge in malaria-endemic countries where a high proportion of children is parasitaemic at any time. A positive malaria blood smear is therefore not specific for severe malaria, and neither are clinical signs, which are similar to those of other severe childhood infections [1]–[3]. Overdiagnosis of falciparum malaria in severely ill children is an important problem in sub-Saharan Africa [4],[5]. Misdiagnosis is associated with increased mortality [6]. Autopsy studies in children dying with “slide-positive” cerebral malaria show an alternative diagnosis in up to 23% of cases [4]. The central pathological process in severe falciparum malaria is sequestration of trophozoite - and schizont-stage–infected erythrocytes in venules and capillaries, which compromise microcirculatory flow to vital organs [7]. The circulating young ring-form parasites do not sequester and therefore do not reflect accurately the sequestered parasite burden. Thus peripheral parasite counts have weak prognostic significance [8],[9], although this can be improved by assessing the stage of development of these peripheral blood parasites or counting the numbers of malaria pigment–containing neutrophils, which reflects recent schizogony [10],[11].
Plasmodium falciparum histidine-rich protein 2 (PfHRP2) is a water-soluble protein found inside the malaria parasite and host erythrocyte, and that circulates free or bound to proteins or antibodies in the plasma compartment [12],[13]. PfHRP2 production peaks during the trophozoite stage, and approximately 90% is released during schizont rupture [14]. Since released PfHRP2 is distributed through the total plasma volume, plasma PfHRP2 can be considered a measure of total parasite burden of the preceding 48-hour asexual parasite life cycle [14],[15]. Studies in Asian adults have shown a strong correlation between plasma PfHRP2, disease severity, and outcome [15],[16].
In the current study we assessed the prognostic significance of plasma PfHRP2 in African children with severe malaria and tested the hypothesis that its assessment could distinguish children with “true” severe malaria, in need of urgent antimalarial treatment, from those with non-malarial severe febrile illness and coincidental peripheral blood parasitaemia, in whom alternative diagnoses and additional treatment need to be considered.
Methods
The study was part of a large multinational trial comparing quinine and artesunate for the treatment of severe malaria in African children (“AQUAMAT,” ISRCTN 50258054), undertaken between October 2005 and July 2010 [17]. Ethics approval was granted by the Oxford Tropical Research Ethics Committee and the countries' ethics review boards. Full details of this trial have been described elsewhere [17]. In brief, children with signs of severe malaria confirmed by a positive P. falciparum lactate dehydrogenase (pLDH)-based rapid diagnostic test were included, provided their parents or caregivers gave full written informed consent. Severity was defined by clinical criteria (see Text S1). Patients were excluded if treated parenterally for >24 hours before admission. Patients were randomised to treatment with either parenteral artesunate or quinine. A venous blood sample was taken for peripheral blood slide, haematocrit (Hct), PfHRP2, biochemistry, and acid-base parameters (EC8+ cartridge for the i-STAT handheld blood analyser). Slide reading was performed by expert microscopists at the Mahidol-Oxford Tropical Medicine Research Unit, and parasites/µl was calculated from thin film (count/1,000 RBC×125.6×Hct) or thick film (count/200 WBC×40) [18],[19].
Plasma PfHRP2 was assessed blinded to patient outcomes from freeze-thawed EDTA plasma samples by a commercial sandwich ELISA kit (Celisa, Cellabs, Sydney, Australia), according to the manufacturer's instructions with minor modifications [15]. Pooled reference plasma from 20 subjects with P. falciparum parasitaemia >200,000/µl was calibrated with recombinant PfHRP2 standard (kindly provided by D. Sullivan, John Hopkins School of Public Health, Baltimore, Maryland, US) and used to construct standard curves. Concentrations in duplicate plasma dilutions (1/25 to 1/3,125 in PBS/0.01%Tween) were determined according to the linear segment of the standard curve, with re-assay in cases where duplicates differed by more than 50%. Plasma samples for PfHRP2 were received from nine of the 11 “AQUAMAT” research sites in seven countries (Mozambique, The Gambia, Kenya, Tanzania, Uganda, Rwanda, and the Democratic Republic of the Congo). The study site in Ghana did not collect samples, and the samples from Nigeria defrosted during transportation.
Individual Patient Estimation of Parasite Burden
Estimation of the total body parasite burden from plasma PfHRP2 has been described in detail in Asian adults with severe malaria and requires incorporation of an elimination half-life estimate [15]. This was assessed separately in African children because clearance might be dependent on immunity (antibodies against PfHRP2), which has a higher level in high transmission settings, and PfHRP2 production is parasite strain dependent [20]. Plasma PfHRP2 half-life was assessed in 30 patients from Tanzania from samples taken on admission and after 3 and 7 days following treatment. Separate ethical approval for this sub-study was obtained from the Ethics Committee of the National Institute for Medical Research, Tanzania. These data were analysed using WinNonlin statistical package (Pharsight, Mountain View, California, US). Individual PfHRP2 concentration-time curves were fitted according to a first-order elimination model. From this, a mean (95%CI) plasma elimination half-life (t½) was estimated as 1.10 (0.91 to 1.29) days, or 0.55 erythrocytic cycles. Half-life was not significantly different between treatment arms, and was not correlated with renal function (estimated by blood urea nitrogen [BUN]). A parasite multiplication factor of 3 immediately before peak parasitaemia was assumed, based on in-vitro and Saimiri monkey studies of African parasite strains causing severe malaria [21],[22]. Higher multiplication rates were explored in a sensitivity analysis [23],[24]. The formula for total parasite burden is: Ptot = 7.3×PfHRP2×(1−Hct)×body weight [kg] ×1013, with PfHRP2 in g/L [15]. The differences in the current formula with the one used earlier in adult Asian patients result from the different estimates for plasma PfHRP2 half-life and parasite multiplication rates. The circulating parasite burden was calculated from the peripheral blood: parasites/µl×106×blood volume ( = 0.08×weight [kg]) [15]. The sequestration index was calculated as total parasite burden/circulating burden [25].
Statistical Analysis
Data were analysed with STATA, version 10 (Stata Corp., Texas, US). Categorical variables were compared between survivors and fatal cases with Chi-squared or Fisher's exact test. Normally distributed or log10-normalized variables were compared using a Student's t-test, the remainder by Wilcoxon rank-sum test. For lowest, middle, and highest tertiles of plasma PfHRP2, comparisons were made between peripheral blood parasitaemia, sequestration index, and treatment effect (mortality) following artesunate versus quinine treatment.
To determine the prognostic significance of plasma PfHRP2, a logistic regression model was constructed with in-hospital death as the dependent variable and PfHRP2 as the independent variable. Since the risk of death showed a non-linear association with log10 PfHRP2 (Figure 1, top), both first - and second-degree fractional polynomial functions were explored to find the optimal fit. A quadratic polynomial function provided the best fit using the likelihood ratio test and by comparison of AUCs (areas under the curve). The regression model was stratified for study site and adjusted for treatment and other established predictors of death, including coma, convulsions, prostration, hypoglycaemia, respiratory distress, shock (combined compensated and decompensated), parasitaemia (/µL), haemoglobin (Hb; g/dL), blood urea nitrogen (BUN; mg/dL), and base excess (BE; mmol/L) [8],[9]. Using a stepwise approach, only covariates that were significant at p<0.01 were retained in the final model. Fit of the final logistic regression model was confirmed using the Hosmer-Lemeshow goodness-of-fit test after ordering the data on predicted probabilities and then regrouping the data into 10 nearly equal-sized groups [26].

Any interaction with transmission intensity regarding associations between plasma PfHRP2 and survival was checked and accounted for if significant. Study sites in Mozambique and The Gambia were defined as low transmission; Rwanda, Tanzania, and Kenya as intermediate; and study sites in Uganda and the Democratic Republic of Congo as high transmission.
Modelling Malaria-Attributable Mortality Based on Plasma PfHRP2
A mechanistic model was constructed to describe the observed U-shaped relationship between PfHRP2 strata and probability of in-hospital death (Figure 1, top), making the following assumptions: (1) an exponential increase of malaria-attributable mortality with plasma PfHRP2, which describes the right side of the curve in Figure 1 (top): Prdeath|malaria = −1+exp(k1logPfHRP2k2); (2) a probability of severe febrile illness due to non-malaria, which decreased exponentially with increasing logPfHRP2: Prnon-malaria = exp(−k4logPfHRP2); (3) a risk of death in patients with non-malaria infection equal to 0.3, independent of plasma PfHRP2: Prdeath|non-malaria = 0.3 [5],[27]; and (4) that 20% of all deaths were due to non-malaria illness: Deathnon-malaria/Deathtotal = 0.2 [4]. The number of non-malarial deaths according to PfHRP2 stratum is then given by:
Deathnon-malaria = Prdeath|non-malaria×Prnon-malaria×Casestotal and the number of deaths due to malaria by Deathmalaria = Prdeath|malaria×(1−Prnon-malaria)×Casestotal. For more details, see Text S2. The effects of assumptions 3 and 4 were explored in a sensitivity analysis.
Results
Patient Characteristics
Of the 5,425 children with pLDH-based rapid diagnostic test RDT confirmed falciparum malaria included in the “AQUAMAT” trial, plasma PfHRP2 was measured in 3,826 patients. PfHRP2 could not be measured in 1,600 (29%) patients because the sample was either not collected or not received in optimal condition. Patients without PfHRP2 data did not differ from the remainder regarding malaria slide positivity rate, geometric mean parasitaemia, or case fatality rate. Baseline clinical and laboratory characteristics according to outcome are summarized in Table 1. Although many clinical and laboratory variables associated with severity differed between survivors and fatal cases, admission parasitaemia did not.

Plasma PfHRP2 in Relation to Disease Severity
PfHRP2 was detectable in 3,800/3,826 (99%) patients with severe malaria. A detectable plasma PfHRP2 (geometric mean, 95% CI, 450 ng/mL, 209 to 966 ng/mL) with a negative blood slide result (but positive malaria RDT) was found in 36 (0.9%) children. Geometric mean plasma PfHRP2 (95%CI) in survivors was 1,046 ng/mL (991 to 1,104 ng/mL) versus 1,611 ng/mL (1,350 to 1,922 ng/mL) in fatal cases (p<0.0001, Table 2). There was no heterogeneity by stratification for transmission intensity in the difference of plasma PfHRP2 concentrations between survivors and fatal cases (p = 0.1).

Plasma PfHRP2 concentrations in relation to established features of severe falciparum malaria are summarized in Table 2. Plasma PfHRP2 was significantly higher in patients with coma, acidosis, and severe anaemia but not in those with shock.
Estimated Total Body Parasite Burden
Geometric mean (95% CI) PfHRP2-derived total parasite burden was 7.5×1011 (7.2×1011 to 7.9×1011) parasites/body (n = 3,800); this was greater in fatal cases (1.2×1012 [1.0×1012 to 1.5×1012], n = 327) than in survivors (7.2×1011 [6.8×1011 to 7.6×1011], n = 3,070, p<0.0001) (Figure 2). In contrast, the total circulating peripheral blood parasite burden did not differ significantly between survivors and fatal cases (p = 0.66). The geometric mean (95%CI) calculated sequestration index, the ratio of total parasitaemia to circulating parasitaemia was 17 (15 to 18) in survivors, versus 30 (23 to 40) in fatal cases (p = 0.0001). The sequestered parasite burden, calculated by subtracting the circulating parasite burden from the total parasite burden, gave a negative result in 296/3,397 (8.7%) patients. Excluding these patients, the geometric mean (95%CI) total sequestered parasite burden was 7.7×1011 parasites/body (7.3×1011 to 8.2×1011, n = 3,101). A sensitivity analysis varying the multiplication factor and PfHRP2 plasma half-life is shown in Text S3.

Plasma PfHRP2 and Risk of Death
There was a U-shaped association between plasma PfHRP2 and risk of death with a nadir in case fatality rate at a logPfHRP2 of 2.24 ( = 174 ng/mL; Figure 1, top). In an adjusted logistic regression model, stratified by study site, plasma PfHRP2 was a strong independent predictor of death. Odds for death were 20% higher per unit increase in logPfHRP2 (adjusted odds ratio [AOR] 1.21, 95%CI 1.05 to 1.39, p = 0.009) above a threshold logPfHRP2 value of 2.24 ( = 174 ng/mL). Below this concentration, risk of death increased with decreasing plasma logPfHRP2 (AOR 2.3, 95%CI 1.1 to 5.0; p = 0.03). The final model was adjusted for plasma BE, BUN, coma, convulsions, hypoglycaemia, peripheral blood parasitaemia, and antimalarial treatment (Hosmer-Lemeshow p-value for goodness-of-fit = 0.35).
Distinguishing Death Attributable to Severe Malaria from Death Attributable to Other Causes
High mortality rates were associated with either very low or very high values of plasma PfHRP2 (Figure 1, top), with the former presumably resulting from a disease other than malaria (including sepsis). The observed case fatalities in the lowest PfHRP2 half log stratum and the higher PfHRP2 strata of ≥3.5–4.0 were both over 15%. A mechanistic model describing the U-shaped correlation between logPfHRP2 stratum and risk of death showed a good fit with the observed data and the statistical model (Figure 1, top). This model was deconvoluted into two separate functions corresponding to non-malaria - and malaria-attributable case fatality rates (Figure 1, bottom). The model showed that below a plasma logPfHRP2 value of 2.24 ( = 174 ng/mL) (derived from the nadir in the polynomial logistic regression model), the probability that death resulted from malaria fell below 50%, corresponding to overall proportions of malaria-attributable severe disease <90% (Figure 1, bottom). In the logPfHRP2 stratum of 3 to 3.5 (1,000 to 3,162 ng/mL) and above, the absolute risk of death due to malaria exceeded 8% with a probability of “true” severe malaria >95% and a probability that a death was caused by severe malaria >85% (Figure 1, bottom). For a sensitivity analysis of the mechanistic model see Text S2.
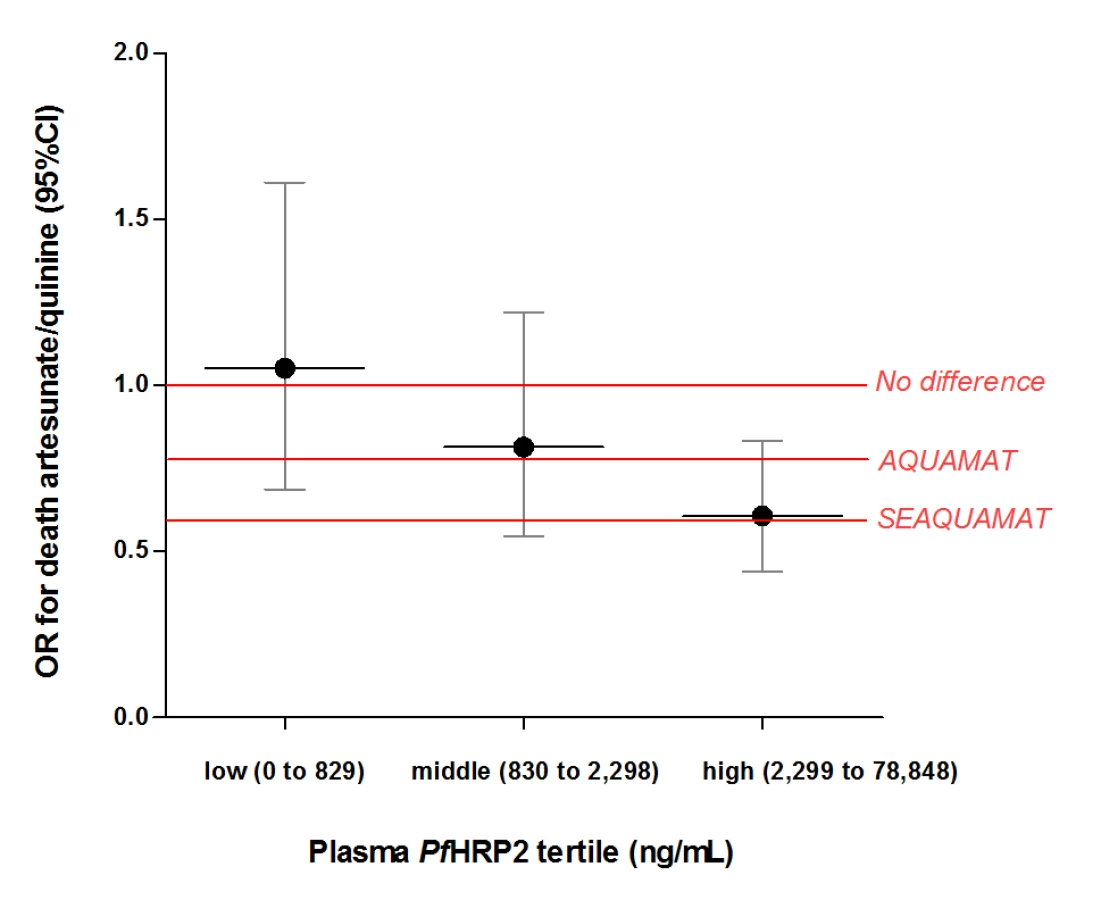
In patients within the highest PfHRP2 tertile, corresponding to logPfHRP2 ≥3.4 (2,300 ng/mL), the odds ratio (OR) for death in patients treated with artesunate versus quinine was 0.61 (95%CI 0.44 to 0.83, p = 0.0018). In patients in the lowest PfHRP2 tertile, there was no difference in mortalities with an OR for death of 1.05 (95%CI 0.69 to 1.61; p = 0.82, Figure 3). The geometric mean (95%CI) sequestration index, the ratio of total to circulating parasite numbers was 69.8 (60.8 to 80.1) in patients in the highest and 4.6 (4.0 to 5.3) in the lowest PfHRP2 tertile (Table 3).

Discussion
This very large prospective study in African children with severe falciparum malaria shows the strong and independent prognostic value of admission plasma PfHRP2 concentration, but not the conventional peripheral blood malaria parasite count. In addition, plasma PfHRP2 was found to be the best immediate measure available to distinguish severe disease caused by malaria from severe febrile illness resulting from another disease with incidental P. falciparum parasitaemia. Since PfHRP2 is a measure of total parasite burden, this suggest a critical pathophysiological role played by sequestered parasites in severe falciparum malaria. This is supported by studies which have correlated obstruction of microcirculatory flow in the rectal and retinal circulations to disease severity and outcome, the strong prognostic value of metabolic acidosis in severe malaria, and autopsy studies showing intense sequestration in vital organs [8],[25],[28]–[31].
These results suggest that in areas of moderate or high malaria transmission where a high proportion of children are parasitaemic, admission plasma PfHRP2 can differentiate children at highest risk of death due to severe falciparum malaria from those with likely alternative causes of severe febrile illness. These findings are supported by several observations.
Firstly, plasma PfHRP2 derived total parasite numbers (geometric mean 7.5×1011/body) are biologically plausible, and were significantly higher in fatal cases. In contrast, less pathogenic circulating peripheral blood parasite numbers were not correlated with a fatal outcome. The calculated sequestration index was 17 in surviving patients and 30 in non-survivors, which is similar to the median (IQR) sequestration index of 40 (9.9–273.8) calculated directly from post-mortem blood vessel counts in 50 Thai and Vietnamese adults who died from cerebral malaria [25].
Second, the U-shaped curve with a nadir at 174 ng/mL describing the relationship between PfHRP2 and risk of death fits with the assumption that with low PfHRP2, death is caused by non-malarial febrile illnesses (including sepsis) which are independent of the low parasite burden, whereas in patients with plasma PfHRP2 above this nadir the probability of death increases with PfHRP2, representing “true” severe malaria with increasing sequestered parasite burdens. The mechanistic model based on these assumptions had a close fit with the observed data. An alternative explanation could be the presence of highly virulent parasite strains causing severe disease independent of a high total parasite burden. However, this would result in a PfHRP2-independent mortality at the left side of the curve and cannot explain the U shape that was actually observed. Assumptions in constructing the mechanistic model included an alternative cause of death in 20% of patients and a risk of death in non-malaria disease of 30%, based on published autopsy and clinical microbiology data [4],[27]. However, the conclusions were not dependent on these assumptions and were robust to the plausible ranges of values defined for the sensitivity analysis.
Third, the treatment benefit of artesunate over quinine was absent in patients in the lowest PfHRP2 tertile, and strongest in the highest tertile (OR 0.61, 95%CI 0.44 to 0.83, p = 0.0018). Since injectable artesunate can benefit only patients with “true” severe malaria, this provides strong supportive evidence that patients with high PfHRP2 do represent this group, and patients with low PfHRP2 do not. The OR of 0.61 in the highest PfHRP2 tertile is remarkably close to the OR of 0.60 (95%CI 0.45 to 0.79) reported in the large SEAQUAMAT trial comparing artesunate with quinine in the treatment of severe falciparum malaria in 1,461 patients in low-transmission settings in Asia [32]. In these epidemiological settings incidental peripheral blood malaria parasitaemia is rare. The diagnosis of severe malaria based on a peripheral blood slide is therefore highly specific, and so the treatment effect of artesunate over quinine is undiluted by non-malarial disease.
Identification of children with slide-positive severe febrile illness but who do not have severe malaria is important for patient management, since overdiagnosis of severe malaria is associated with increased mortality [6]. A low plasma PfHRP2 should prompt investigation of alternative diagnoses including septicaemia, early administration of parenteral broad spectrum antibiotics (if not already routine), and intensive monitoring. Often antibiotics are given only after a disappointing clinical response to antimalarials, which may be too late. High plasma PfHRP2 concentrations should not discourage antibiotic treatment combined with antimalarial treatment, because of the high proportion of concomitant invasive bacterial disease [2]. Patients with high plasma PfHRP2, which indicates “true” severe malaria with a poor prognosis, should be monitored closely, preferentially in a high-dependency or intensive care unit. As a tool in the design of clinical trials, plasma PfHRP2 is substantially better than peripheral blood parasitaemia in assessing the malaria-attributable fractions and defining the group of patients with “true” severe malaria and a high risk of death (Figure S3 in Text S4 and [33]) An alternative tool is the presence of malaria retinopathy, which has been shown to be highly specific for cerebral malaria as confirmed by post-mortem autopsy [4], although this tool does require training and skilled ophthalmoscopy [34],[35]. It has been evaluated for cerebral malaria [36],[37], whereas many patients with severe falciparum malaria present with other syndromes [17]. PfHRP2 can be used in both cerebral and severe non-cerebral malaria. A direct comparison between the two methods is currently underway. Development of a semi-quantitative rapid test for the detection of plasma PfHRP2 with carefully chosen thresholds could be a valuable tool in high transmission settings to distinguish “true” severe malaria from severe non-malarial febrile illness. For example, a plasma PfHRP2 concentration of >1,000 ng/mL (62.1% of cases in our cohort) denotes a probability >95% of “true” severe malaria with an overall case fatality rate of 11.6% (95%CI 10.3 to 12.9). Defining populations with “true” severe malaria and high mortality is thus critical information for clinicians as well as researchers. In contrast, a plasma PfHRP2 concentration <100 ng/mL (8.1% of cases in our cohort) denotes a probability >15% that severe non-malarial illness is the cause of illness, warranting additional investigations.
Limitations of this study include the inherent dependency of the models on certain assumptions. Estimating the total parasite burden from PfHRP2 is sensitive to the assumed parasite multiplication factor. In the current study the multiplication rate was assumed to be 3, based on in vitro data comparing multiplication rates and multiplication potency of parasites obtained from African children compared to Asian adults. The multiplication rate of 8 used in the original model in Asian adults was based on non-immune adult patient data from the era of malaria therapy of neurosyphilis, and comparable information is obviously not available for our patient group. Applying this higher multiplication rate in this study results in an implausibly high estimated total parasite burden. In addition to differences in parasite multiplication rates, the calculated total parasite burden is dependent on the assumed half-life of plasma PfHRP2, which can vary between patients, and on the amount of PfHRP2 released per parasite per cycle, which can vary between strains [15],[20]. A sensitivity analysis of these parameters is shown in Figure S2 in Text S3. The half-life of plasma PfHRP2 in the current study was shorter than observed in adult patients in Southeast Asia (mean 1.1 versus 3.7 days) [15]. This is presumably related to the African setting where malaria transmission is high and immunological factors including high PfHRP2 antibody titres could increase plasma clearance of PfHRP2 [38],[39]. Since variations in the model parameter estimates are applied to the entire patient group, the model renders either pathophysiologically implausible upper (more parasites than the number of circulating red cells) or lower limits (fewer total parasites than the calculated circulating parasitaemia). Actual total parasite numbers can thus be slightly different from the model estimates. However, differences in the calculated total parasite burdens between subgroups do not depend on the choice of these variables, since these variables will affect this value by the same factor in all subgroups. A recent study in Papuan children with falciparum malaria did not show a correlation between PfHRP2 and disease severity [40]. However, children (n = 220) in this study diagnosed with severe malaria appeared to be only moderately ill as reflected by the <1% case fatality rate and low plasma PfHRP2 values (median 456 ng/mL), whereas patients in that study considered to have uncomplicated malaria had lower plasma bicarbonate concentrations as a measure of acidosis than those with severe malaria. In the present study, <1% cases had undetectable plasma PfHRP2 concentrations, despite presence of P. falciparum on the blood slide. This could have been caused by genetic variation in PfHRP2 [41], although this polymorphism is thought not to affect the detection by ELISA [42],[43]. Deletions of the PfHRP2 gene have been reported in field isolates from the Amazon region and in a single report from sub-Saharan Africa [44],[45]. However, the incidence of this genotype is thought to be low in parasites causing severe malaria related to reduced parasite fitness [46],[47]. A study sequencing the PfHRP2 gene in parasites from all patients in the current study who had low plasma PfHRP2 concentrations is underway.
In conclusion, admission plasma PfHRP2 provides a tool in areas of moderate and high malaria transmission to distinguish “true” severe falciparum malaria from severe febrile illness with incidental malaria parasitaemia. Plasma PfHRP2 concentrations are a valuable prognosticator in African children with severe falciparum malaria.
Supporting Information
Zdroje
1. EnglishM, PuntJ, MwangiI, McHughK, MarshK (1996) Clinical overlap between malaria and severe pneumonia in Africa children in hospital. Trans R Soc Trop Med Hyg 90 : 658–662.
2. BerkleyJ, MwarumbaS, BramhamK, LoweB, MarshK (1999) Bacteraemia complicating severe malaria in children. Trans R Soc Trop Med 93 : 283–286.
3. EvansJA, AduseiA, TimmannC, MayJ, MackD, et al. (2004) High mortality of infant bacteraemia clinically indistinguishable from severe malaria. QJM 97 : 591–597.
4. TaylorTE, FuWJ, CarrRA, WhittenRO, MuellerJS, et al. (2004) Differentiating the pathologies of cerebral malaria by postmortem parasite counts. Nat Med 10 : 143–145.
5. BerkleyJA, LoweBS, MwangiI, WilliamsT, BauniE, et al. (2005) Bacteremia among children admitted to a rural hospital in Kenya. N Engl J Med 352 : 39–47.
6. ReyburnH, MbatiaR, DrakeleyC, CarneiroI, MwakasungulaE, et al. (2004) Overdiagnosis of malaria in patients with severe febrile illness in Tanzania: a prospective study. BMJ 329 : 1212.
7. DondorpAM, PongponratnE, WhiteNJ (2004) Reduced microcirculatory flow in severe falciparum malaria: pathophysiology and electron-microscopic pathology. Acta Trop 89 : 309–317.
8. MarshK, ForsterD, WaruiruC, MwangiI, WinstanleyM, et al. (1995) Indicators of life-threatening malaria in African children. N Engl J Med 332 : 1399–1404.
9. JaffarS, Van HensbroekMB, PalmerA, SchneiderG, GreenwoodB (1997) Predictors of a fatal outcome following childhood cerebral malaria. Am J Trop Med Hyg 57 : 20–24.
10. SilamutK, WhiteNJ (1993) Relation of the stage of parasite development in the peripheral blood to prognosis in severe falciparum malaria. Trans R Soc Trop Med Hyg 87 : 436–443.
11. NguyenPH, DayN, PramTD, FergusonDJ, WhiteNJ (1995) Intraleucocytic malaria pigment and prognosis in severe malaria. Trans R Soc Trop Med Hyg 89 : 200–204.
12. HowardRJ, UniS, AikawaM, AleySB, LeechJH, et al. (1986) Secretion of a malarial histidine-rich protein (Pf HRP II) from Plasmodium falciparum-infected erythrocytes. J Cell Biol 103 : 1269–1277.
13. KifudeCM, RajasekariahHG, SullivanDJJr, StewartVA, AngovE, et al. (2008) Enzyme-linked immunosorbent assay for detection of Plasmodium falciparum histidine-rich protein 2 in blood, plasma, and serum. Clin Vaccine Immunol 15 : 1012–1018.
14. DesakornV, DondorpAM, SilamutK, PongtavornpinyoW, SahassanandaD, et al. (2005) Stage-dependent production and release of histidine-rich protein 2 by Plasmodium falciparum. Trans R Soc Trop Med Hyg 99 : 517–524.
15. DondorpAM, DesakornV, PongtavornpinyoW, SahassanandaD, SilamutK, et al. (2005) Estimation of the total parasite biomass in acute falciparum malaria from plasma PfHRP2. PLoS Med 2: e204 doi:10.1371/journal.pmed.0020204.
16. YeoTW, LampahDA, GitawatiR, TjitraE, KenangalemE, et al. (2008) Angiopoietin-2 is associated with decreased endothelial nitric oxide and poor clinical outcome in severe falciparum malaria. Proc Natl Acad Sci USA 105 : 17097–17102.
17. DondorpAM, FanelloCI, HendriksenIC, GomesE, SeniA, et al. (2010) Artesunate versus quinine in the treatment of severe falciparum malaria in African children (AQUAMAT): an open-label, randomised trial. Lancet 376 : 1647–1657.
18. GreenwoodBM, ArmstrongJR (1991) Comparison of two simple methods for determining malaria parasite density. Trans R Soc Trop Med Hyg 85 : 186–188.
19. PlancheT, KrishnaS, KombilaM, EngelK, FaucherJF, et al. (2001) Comparison of methods for the rapid laboratory assessment of children with malaria. Am J Trop Med Hyg 65 : 599–602.
20. BakerJ, GattonML, PetersJ, HoMF, McCarthyJS, et al. (2011) Transcription and expression of Plasmodium falciparum histidine-rich proteins in different stages and strains: implications for rapid diagnostic tests. PLoS One 6: e22593 doi:10.1371/journal.pone.0022593.
21. DeansAM, LykeKE, TheraMA, PloweCV, KoneA, et al. (2006) Low multiplication rates of African Plasmodium falciparum isolates and lack of association of multiplication rate and red blood cell selectivity with malaria virulence. Am J Trop Med Hyg 74 : 554–563.
22. Le ScanfC, Vigan-WomasI, ContaminH, GuillotteM, BischoffE, et al. (2008) Rosetting is associated with increased Plasmodium falciparum in vivo multiplication rate in the Saimiri sciureus monkey. Microbes Infect 10 : 447–451.
23. ChotivanichK, UdomsangpetchR, SimpsonJA, NewtonP, PukrittayakameeS, et al. (2000) Parasite multiplication potential and the severity of Falciparum malaria. J Infect Dis 181 : 1206–1209.
24. SimpsonJA, AaronsL, CollinsWE, JefferyGM, WhiteNJ (2002) Population dynamics of untreated Plasmodium falciparum malaria within the adult human host during the expansion phase of the infection. Parasitology 124 : 247–263.
25. SilamutK, PhuNH, WhittyC, TurnerGD, LouwrierK, et al. (1999) A quantitative analysis of the microvascular sequestration of malaria parasites in the human brain. Am J Pathol 155 : 395–410.
26. StataCorp (2007) Stata 10 Base Reference Manual (L): College Station, TX: Stata Press.
27. BerkleyJA, BejonP, MwangiT, GwerS, MaitlandK, et al. (2009) HIV infection, malnutrition, and invasive bacterial infection among children with severe malaria. Clin Infect Dis 49 : 336–343.
28. DondorpAM, InceC, CharunwatthanaP, HansonJ, van KuijenA, et al. (2008) Direct in vivo assessment of microcirculatory dysfunction in severe falciparum malaria. J Infect Dis 197 : 79–84.
29. WhiteVA, LewallenS, BeareNA, MolyneuxME, TaylorTE (2009) Retinal pathology of pediatric cerebral malaria in Malawi. PLoS One 4: e4317 doi:10.1371/journal.pone.0004317.
30. BeareNA, HardingSP, TaylorTE, LewallenS, MolyneuxME (2009) Perfusion abnormalities in children with cerebral malaria and malarial retinopathy. J Infect Dis 199 : 263–271.
31. MacPhersonGG, WarrellMJ, WhiteNJ, LooareesuwanS, WarrellDA (1985) Human cerebral malaria. A quantitative ultrastructural analysis of parasitized erythrocyte sequestration. Am J Pathol 119 : 385–401.
32. DondorpA, NostenF, StepniewskaK, DayN, WhiteN (2005) Artesunate versus quinine for treatment of severe falciparum malaria: a randomised trial. Lancet 366 : 717–725.
33. BejonP, BerkleyJA, MwangiT, OgadaE, MwangiI, et al. (2007) Defining childhood severe falciparum malaria for intervention studies. PLoS Med 4: e251 doi: 10.1371/journal.pmed.0040251.
34. BurtonM, Nyong'oO, BurtonK, JohnW, InkoomE, et al. (2004) Retinopathy in Gambian children admitted to hospital with malaria. Trop Doct 34 : 214–218.
35. BeareNA, TaylorTE, HardingSP, LewallenS, MolyneuxME (2006) Malarial retinopathy: a newly established diagnostic sign in severe malaria. Am J Trop Med Hyg 75 : 790–797.
36. EssumanVA, Ntim-AmponsahCT, AstrupBS, AdjeiGO, KurtzhalsJA, et al. (2010) Retinopathy in severe malaria in Ghanaian children–overlap between fundus changes in cerebral and non-cerebral malaria. Malar J 9 : 232.
37. BeareNA, SouthernC, ChaliraC, TaylorTE, MolyneuxME, et al. (2004) Prognostic significance and course of retinopathy in children with severe malaria. Arch Ophthalmol 122 : 1141–1147.
38. BiswasS, TomarD, RaoDN (2005) Investigation of the kinetics of histidine-rich protein 2 and of the antibody responses to this antigen, in a group of malaria patients from India. Ann Trop Med Parasitol 99 : 553–562.
39. AbekuTA, KristanM, JonesC, BeardJ, MuellerDH, et al. (2008) Determinants of the accuracy of rapid diagnostic tests in malaria case management: evidence from low and moderate transmission settings in the East African highlands. Malar J 7 : 202.
40. ManningL, LamanM, StanisicD, Rosanas-UrgellA, BonaC, et al. (2011) Plasma Plasmodium falciparum histidine-rich protein-2 concentrations do not reflect severity of malaria in Papua new guinean children. Clin Infect Dis 52 : 440–446.
41. BakerJ, McCarthyJ, GattonM, KyleDE, BelizarioV, et al. (2005) Genetic diversity of Plasmodium falciparum histidine-rich protein 2 (PfHRP2) and its effect on the performance of PfHRP2-based rapid diagnostic tests. J Infect Dis 192 : 870–877.
42. MartinSK, RajasekariahGH, AwindaG, WaitumbiJ, KifudeC (2009) Unified parasite lactate dehydrogenase and histidine-rich protein ELISA for quantification of Plasmodium falciparum. Am J Trop Med Hyg 80 : 516–522.
43. BakerJ, HoMF, PelecanosA, GattonM, ChenN, et al. (2010) Global sequence variation in the histidine-rich proteins 2 and 3 of Plasmodium falciparum: implications for the performance of malaria rapid diagnostic tests. Malar J 9 : 129.
44. HouzeS, HubertV, Le PessecG, Le BrasJ, ClainJ (2011) Combined deletions of pfhrp2 and pfhrp3 genes result in Plasmodium falciparum malaria false-negative rapid diagnostic test. J Clin Microbiol 49 : 2694–2696.
45. KoitaOA, DoumboOK, OuattaraA, TallLK, KonareA, et al. (2012) False-negative rapid diagnostic tests for malaria and deletion of the histidine-rich repeat region of the hrp2 gene. Am J Trop Med Hyg 86 : 194–198.
46. WellemsTE, WallikerD, SmithCL, do RosarioVE, MaloyWL, et al. (1987) A histidine-rich protein gene marks a linkage group favored strongly in a genetic cross of Plasmodium falciparum. Cell 49 : 633–642.
47. Walker-JonahA, DolanSA, GwadzRW, PantonLJ, WellemsTE (1992) An RFLP map of the Plasmodium falciparum genome, recombination rates and favored linkage groups in a genetic cross. Mol Biochem Parasitol 51 : 313–320.
Štítky
Interní lékařstvíČlánek vyšel v časopise
PLOS Medicine
2012 Číslo 8
- Alternativní léčebné možnosti u hypercholesterolemie při intoleranci statinů
- Vliv kombinace nutraceutik na remodelaci levé komory srdeční u osob s metabolickým syndromem
- Nutraceutika a jejich ovlivnění mírného kardiometabolického rizika
- Princip účinku medu v léčbě chronických i infikovaných ran
- Superoxidovaný roztok a jeho využití v léčbě ran
Nejčtenější v tomto čísle
- Feasibility, Yield, and Cost of Active Tuberculosis Case Finding Linked to a Mobile HIV Service in Cape Town, South Africa: A Cross-sectional Study
- Multidrug Resistant Pulmonary Tuberculosis Treatment Regimens and Patient Outcomes: An Individual Patient Data Meta-analysis of 9,153 Patients
- Child Mortality Estimation: A Global Overview of Infant and Child Mortality Age Patterns in Light of New Empirical Data
- What Is the Optimal First Line Antiretroviral Therapy in Resource-Limited Settings?