
IscR Is Essential for Type III Secretion and Virulence
Bacterial pathogens use regulators that sense environmental cues to enhance their fitness. Here, we identify a transcriptional regulator in the human gut pathogen, Yersinia pseudotuberculosis, which controls a specialized secretion system essential for bacterial growth in mammalian tissues. This regulator was shown in other bacterial species to alter its activity in response to changes in iron concentration and oxidative stress, but has never been studied in Yersinia. Importantly, Y. pseudotuberculosis experiences large changes in iron bioavailability upon transit from the gut to deeper tissues and iron is a critical component in Yersinia virulence, as individuals with iron overload disorders have enhanced susceptibility to systemic Yersinia infections. Our work places this iron-modulated transcriptional regulator within the regulatory network that controls virulence gene expression in Y. pseudotuberculosis, identifying it as a potential new target for antimicrobial agents.
Published in the journal:
. PLoS Pathog 10(6): e32767. doi:10.1371/journal.ppat.1004194
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1004194
Summary
Bacterial pathogens use regulators that sense environmental cues to enhance their fitness. Here, we identify a transcriptional regulator in the human gut pathogen, Yersinia pseudotuberculosis, which controls a specialized secretion system essential for bacterial growth in mammalian tissues. This regulator was shown in other bacterial species to alter its activity in response to changes in iron concentration and oxidative stress, but has never been studied in Yersinia. Importantly, Y. pseudotuberculosis experiences large changes in iron bioavailability upon transit from the gut to deeper tissues and iron is a critical component in Yersinia virulence, as individuals with iron overload disorders have enhanced susceptibility to systemic Yersinia infections. Our work places this iron-modulated transcriptional regulator within the regulatory network that controls virulence gene expression in Y. pseudotuberculosis, identifying it as a potential new target for antimicrobial agents.
Introduction
Type III secretion systems (T3SS) are important components in the progression of disease for a number of clinically relevant human pathogens, including those in the genera Shigella, Salmonella, Escherichia, Chlamydia, Vibrio, Pseudomonas, and Yersinia [1], [2]. The T3SS functions as an injectisome that delivers bacterial effector proteins directly into the host cell cytoplasm [2]. While the T3SS apparatus itself is structurally conserved, the repertoire of T3SS effector proteins used by each group of pathogens is distinct [2]. Thus, the effect of the T3SS on the host is unique to the needs of the pathogen [2]. While the T3SS is generally essential for a T3SS-expressing pathogen to cause disease, several aspects of the T3SS may be detrimental to bacterial growth [2]. For example, T3SS components are recognized by the host immune system [3], [4]. In addition, expression of the T3SS is energetically costly and, in some organisms, T3SS induction correlates with growth arrest [5]. Therefore, regulation is essential for proper T3SS function in order to ensure that it occurs only during host cell contact in the appropriate host tissue [2], [6].
Members of the genus Yersinia that utilize a T3SS are important human pathogens: Y. pestis, the causative agent of plague, and the enteropathogens Y. enterocolitica and Y. pseudotuberculosis. The Y. pseudotuberculosis Ysc T3SS is encoded on a 70-kb plasmid termed pYV [7]–[9] and is made up of approximately 25 known proteins comprising three main structures: the basal body, the needle apparatus, and the translocon [10], [11]. The basal body, which displays a high degree of similarity to the flagellar basal body, is made up of rings that span the inner and outer membranes and a rod that traverses the periplasmic space [12]. Basal body associated proteins include YscN, an ATPase that aids in the secretion and translocation of effector proteins [13]. The needle complex, which is thought to act as a molecular channel for effector protein translocation, is a straight hollow appendage approximately 60 nm in length and is made up of helical polymerized subunits of YscF [12]. The translocon is comprised of three proteins: YopD, YopB and LcrV, which are essential for pore formation in the target host membrane and proper translocation of effector proteins YopHEMOJTK to the host cytoplasm [12], [14]. Also encoded on pYV are chaperones important for efficient translocation of a subset of effector proteins [15]. Lastly, several transcriptional and post-transcriptional regulators of the T3SS are found on pYV, including the AraC-like transcriptional regulator LcrF. LcrF is responsible for expression of a number of T3SS structural genes and Yop effectors, specifically the virC and lcrGVH-yopBD operons as well as genes encoding effector Yops, the adhesin YadA, and the lipoprotein YlpA [16]-[22]. LcrF itself is thermoregulated at both the transcriptional and translational levels through the action of the histone-like protein YmoA and a cis-acting RNA thermosensor located on the lcrF transcript, respectively [23], [24]. This enables Yersinia to express T3SS genes at 37°C within the mammalian host, but not at lower temperatures [23], [24]. Importantly, proper LcrF-mediated control of T3SS expression is important for Y. pseudotuberculosis virulence [24].
IscR belongs to the Rrf2 family of winged helix-turn-helix transcription factors [25], [26] and has been studied extensively in E. coli, where its DNA-binding activity is dependent on coordination of an iron-sulfur [2Fe-2S] cluster through three conserved cysteines and a histidine [27]–[30]. E. coli IscR recognizes two distinct DNA motifs, type 1 and type 2, depending on the Fe-S status of the protein [31]. Holo-IscR coordinating an Fe-S cluster binds both type 1 and type 2 motifs, while clusterless apo-IscR recognizes only the type 2 DNA-binding motif [27], [32], [33]. As iron starvation, oxidative stress, and oxygen limitation affect the holo-IscR/apo-IscR ratio, these environmental cues are thought to have a direct effect on gene expression through IscR in E. coli [28]–[30]. For example, holo-IscR represses transcription of the housekeeping iscRSUA-hscBA-fdx Fe-S cluster biogenesis operon [32], [34], while either holo - or apo-IscR promotes transcription of the inducible sufABCDSE Fe-S cluster biogenesis operon [33], [35]. Both pathways function to insert Fe-S clusters onto proteins involved in a range of metabolic processes including electron transfer, substrate binding/activation, iron/sulfur storage, regulation, and enzyme activity [36]. In addition, E. coli IscR is also known to regulate transcription of other Fe-S cluster assembly genes such as erpA (yadR) as well as genes integral to oxidative stress resistance, biofilm formation, and anaerobic respiration [28]–[30], [34]. IscR is widely conserved among bacteria [25] and its regulatory activity is integral to the infectious process of the plant pathogen Erwinia chrysanthemi [37]. Furthermore, IscR plays an important role in the virulence of the human pathogens Pseudomonas aeruginosa through modulation of the catalase katA [38], Burkholderia mallei through resistance to reactive nitrogen species [39], and Vibrio vulnificus through induction of several virulence-associated pathways [39], [40]. While the iron-dependent transcriptional repressor Fur has been shown to control T3SS expression in Salmonella and Shigella [41], [42], IscR has never been linked to regulation of the T3SS in any organism and has not been studied in Yersinia.
In this study, we isolated two independent IscR transposon insertion mutants in a novel screen for Y. pseudotuberculosis genes important for T3SS function. We assessed the impact of iscR deletion on Y. pseudotuberculosis in vitro and in vivo growth, type III secretion, and global gene expression. We found IscR to be essential for full T3SS function and virulence in a mouse model of infection. In addition, we provide evidence that IscR control of the T3SS stems from direct transcriptional regulation of the T3SS master regulator LcrF.
Results
IscR is required for Y. pseudotuberculosis Ysc T3SS function
To identify regulators of the Y. pseudotuberculosis T3SS, we utilized a novel screen to isolate transposon mutants with defects in T3SS function. We previously showed that Y. pseudotuberculosis expressing a functional T3SS induces NFκB activation in HEK293T cells [43], enabling us to use host cell NFκB activation as a readout for T3SS function in Y. pseudotuberculosis transposon mutants. As some T3SS effector proteins inhibit NFκB signaling [44], we performed the screen using a Y. pseudotuberculosis transposon mutant library in a genetic background that lacked the known T3SS effector proteins YopHEMOJT (Δyop6; [43]). We identified several transposon mutants with defects in triggering activation of NFκB in HEK293T cells (L. Kwuan, N. Herrera, H. Ramirez, V. Auerbuch, data not shown), suggesting defective T3SS function. Among these were two strains with unique transposon insertions in YPTB2860 (Figure 1A), encoding a protein with 79% identity to the E. coli iron-sulfur cluster regulator IscR, part of the iscRSUA-hscBA-fdx operon involved in Fe-S cluster biogenesis (Figure 1B). Importantly, the helix-turn-helix DNA binding domain as well as the three cysteines and histidine known to coordinate an iron-sulfur (Fe-S) cluster in E. coli IscR are conserved in all three Yersinia species (Figure 1B). These data indicate that Yersinia IscR may coordinate an Fe-S cluster and, as in E. coli, may regulate gene transcription.

To validate that loss of IscR in Y. pseudotuberculosis leads to T3SS defects, we isolated the two iscR transposon mutants (iscR::Tn1 and iscR::Tn2) from our library and again measured their ability to trigger NFκB activation in HEK293T cells compared to the Δyop6 parental strain and a ΔyscNU T3SS-null mutant [43]. In addition, we constructed an in-frame iscR deletion mutant in the Δyop6 genetic background (Δyop6/ΔiscR) and tested it in this assay. We found that disruption of iscR led to ∼2-fold less NFκB activation relative to the Δyop6 T3SS+ parental strain, although NFκB activation levels were still ∼5-fold higher than a strain with complete lack of T3SS function (ΔyscNU; Figure 2A), suggesting that loss of iscR leads to partial T3SS loss.

To verify further that deletion of iscR leads to alterations in T3SS function, we assessed the ability of the Δyop6/ΔiscR mutant to insert YopBD pores in target host cell membranes by measuring entry of ethidium bromide (EtBr) inside Y. pseudotuberculosis-infected bone marrow derived macrophages [45], [46]. Pore formation by the Δyop6/ΔiscR mutant was decreased by 7-fold (p<0.05) relative to the Δyop6 parental strain, which could be restored upon complementation with plasmid-encoded iscR (Figure 2B). To determine whether loss of iscR affects T3SS function in a wild type genetic background, we constructed an in-frame iscR deletion (ΔiscR) in the wild type Y. pseudotuberculosis IP2666 strain expressing six of the seven known T3SS effector proteins YopHEMOJK [47]. We then visualized the secretome of the ΔiscR mutant relative to wild type. Deletion of iscR led to a dramatic decrease in secretion of T3SS cargo relative to the wild type background, which can be restored upon complementation with plasmid-encoded iscR (Figure 2C). Importantly, this lack of type III secretion did not result from a defect in growth of the mutant, as the ΔiscR mutant actually grew better than wild type bacteria under T3SS-inducing conditions (Figure S1A). This is consistent with a T3SS defect in this strain, as wild type Yersinia display a characteristic growth arrest upon T3SS expression [5], [48], [49]. Collectively, these data demonstrate that Y. pseudotuberculosis IscR is required for proper T3SS function.
IscR is required for full virulence of Y. pseudotuberculosis
Based on the knowledge that the T3SS plays an important role in the virulence of human pathogenic Yersinia, we sought to investigate whether the diminished type III secretion observed in the Y. pseudotuberculosis ΔiscR strain would lead to a reduction in the infectious capacity of this mutant. Mice were orogastrically infected with 2×108 CFU of either the Y. pseudotuberculosis wild type or isogenic ΔiscR mutant strains. At 5 days post-inoculation, mice infected with the ΔiscR mutant displayed significantly decreased colonization of Peyer's patches and mesenteric lymph nodes (MLN) as well as diminished systemic colonization (Figure 3). Specifically, we noted 10 - and 130-fold reductions in CFU recovered from the Peyer's patches and MLNs, respectively, in mice infected with the ΔiscR mutant strain relative to wild type. Notably, we observed a 1000 - to 10,000-fold decrease in bacterial burden in the spleen and liver respectively. The diminished ability of the ΔiscR mutant strain to colonize deep tissue sites is underscored by the fact that bacteria were not detected in seven of the nine livers analyzed. These findings suggest that IscR is essential for Y. pseudotuberculosis virulence in an oral infection model.

IscR deletion leads to global misregulation of gene expression in Y. pseudotuberculosis
To begin to understand the mechanistic contribution of IscR to Y. pseudotuberculosis pathogenesis, we performed high throughput transcriptome sequencing (RNAseq) analysis to determine the Y. pseudotuberculosis genes directly and indirectly controlled by IscR under iron replete, T3SS-inducing conditions. Total RNA was collected from wild type Y. pseudotuberculosis as well as the ΔiscR mutant strain grown in M9 at 37°C for 3 h, a point at which the ΔiscR and wild type strains display comparable growth rates (Figure S1A).
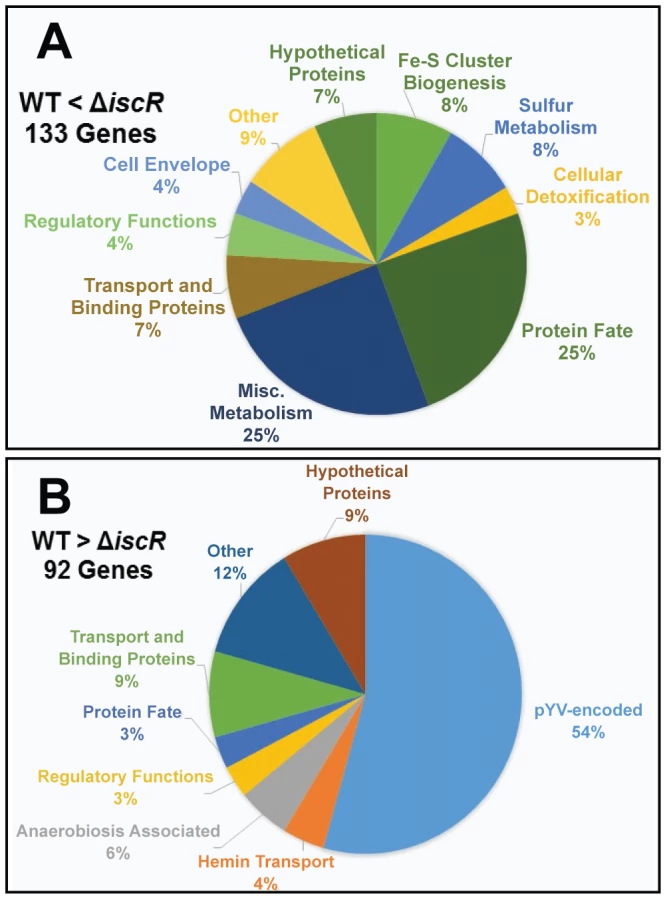
For the ΔiscR mutant relative to the wild type, a total of 226 genes demonstrated a statistically significant fold change of ≥2 (Table S1). Of these, 134 genes were up-regulated in the ΔiscR mutant relative to the wild type (Table 1 & Figure 4A), while 92 were down-regulated (Table 2 & Figure 4B). Genes found to be up-regulated in the ΔiscR mutant include key elements of Fe-S cluster biosynthesis, cellular detoxification, metabolism, and protein fate (Figure 4A). The most notable increases in transcription were observed for genes encoding Fe-S cluster biosynthesis proteins including those encoded in the isc operon, iscS (18.7-fold), iscU (21.7-fold) and iscA (13-fold) (Table 1 & Figure S2A). Additional genes encoding proteins involved in Fe-S cluster assembly and their respective fold increases include iscX/yfhJ (10.8), fdx (10.9), hscB (10), hscA (9.3), yadR/erpA (6.8), pepB (10.1) and nfuA (7.0). To validate these findings, we performed qRT-PCR analysis on the second gene encoded in the iscRSUA operon, iscS, as well as on the gene encoding the Fe-S biosynthesis protein ErpA. Transcription of iscS was increased by 30-fold, while erpA expression was increased 5-fold (p<0.05; Figure 5A). Bioinformatic analysis identified two IscR type 1 motifs upstream of the iscRSUA-hscBA-fdx operon (Figure S2B) as well as one site each located upstream of both erpA and nfuA (data not shown). Based on this data, we propose that Y. pseudotuberculosis IscR modulates Fe-S cluster biosynthesis expression in a manner akin to that of E. coli IscR.



IscR is required for transcription of T3SS genes
In total, 92 genes were significantly down-regulated in the ΔiscR mutant relative to wild type Y. pseudotuberculosis (Table 2). These data demonstrate that the majority of pYV-encoded genes are decreased in the ΔiscR mutant relative to the wild type strain, including genes essential for proper T3SS expression and function. The virC and lcrGVH-yopBD operons as well as genes encoding the T3SS cargo YopHEMOJTK were the most affected upon deletion of iscR: the effector proteins YopJ (−3.4-fold), YopM (−5.3-fold) and YopT (−5.5-fold), the effector protein and translocation regulator YopK (−9.3-fold), as well as a number of genes encoding T3SS structural proteins. Genes encoding regulators that control T3SS expression and function were decreased in the mutant including lcrQ (−2.1-fold), lcrF (−3.3-fold), lcrG (−2.8-fold) and lcrH (−3.9-fold). To verify that T3SS gene expression was indeed decreased in the ΔiscR mutant, we measured the transcript levels of the genes encoding T3SS structural proteins YscN, YscF, and the T3SS transcriptional regulator LcrF via qRT-PCR. As detailed in Figure 5B, we observed fold decreases of 2.8-fold (p<0.05), 6.9-fold (p<0.001), and 5.4-fold (p<0.0001) for yscN, yscF, and lcrF, respectively. These data support our RNAseq analysis and confirm that IscR is required for robust transcription of Y. pseudotuberculosis T3SS genes.
In addition to T3SS genes, 25 other pYV-encoded genes were decreased in the mutant, but these are annotated as hypothetical proteins, transposases, and pseudogenes. Analysis of the relative abundance of pYV in the Y. pseudotuberculosis wild type and ΔiscR strains was performed in order to verify that the decreases in pYV-encoded genes were not a result of plasmid loss (Figure S3). The concentration of plasmid isolated from the wild type and ΔiscR mutant was comparable, suggesting that the decreased transcription of pYV-encoded genes, including those encoding the T3SS, are not a result of decreased stability of the pYV plasmid.
Y. pseudotuberculosis expressing only apo-locked IscR has a proton motive force defect and cannot secrete Yops
To assess the contribution of Fe-S cluster ligation to IscR control of the T3SS, we constructed an IscR mutant strain in which the three conserved cysteines were substituted with alanines (C92A, C98A, C104A; apo-locked IscR). Identical mutations in E. coli IscR render the protein unable to coordinate an iron-sulfur cluster, yet able to bind type 2 DNA binding motifs and to regulate target gene transcription [28]–[30]. We analyzed the secretome of the Y. pseudotuberculosis apo-locked IscR strain under T3SS-inducing conditions and found that the mutant was just as defective as the ΔiscR strain in Yop secretion (Figure 2C). This defect could be complemented with plasmid-encoded wild type IscR. As apo-locked IscR is insufficient to promote type III secretion, holo-IscR-mediated regulation of gene expression through one or more type 1 motifs may be specifically involved in regulating T3SS gene expression. Alternatively, forcing all IscR expression within the cell to the clusterless form, which leads to IscR overexpression, may lead to alterations of bacterial pathways that indirectly affect type III secretion.
Consistent with this latter explanation, the apo-locked IscR mutant exhibited decreased colony size on LB agar, slower growth in rich media (Figure S1), and decreased motility (Figure 6A). The flagellar basal body is a T3SS itself, indicating that the defect in the Ysc T3SS for this strain may be a result of gross abnormalities in secretion systems. Based on these findings, we set out to examine whether the apo-locked IscR mutant demonstrated alterations in membrane potential, as this has been shown to be important for both motility and Ysc T3S in Y. enterocolitica [50]. To this end, we examined bacterial membrane potential under T3SS inducing conditions. As demonstrated in Figure 6B, there is a notable decrease in membrane potential in the apo-locked IscR mutant relative to the wild type strain, which can be complemented upon addition of wild type iscR on a plasmid. Furthermore, the membrane potential of the ΔiscR mutant strain is comparable to that of the wild type. Collectively, these data suggest that the apo-locked IscR mutant has a proton motive force defect, leading to decreased type III secretion and motility. These findings highlight the importance for Yersinia to maintain appropriate levels of holo-IscR relative to apo-IscR in order maintain normal membrane potential.

IscR recognizes a type 2 motif upstream of the yscW-lcrF operon
To begin to understand the nature of the T3SS defect in the presence of only apo-IscR, we carried out RNAseq analysis on the Y. pseudotuberculosis apo-locked IscR mutant grown under T3SS-inducing conditions and compared the results with data from the wild type and ΔiscR strains. Curiously, the apo-locked IscR mutant displayed aberrant expression of genes involved in stress response, transport, cell envelope, as well as electron transport (data not shown). Of note, the Fe-S cluster biosynthesis proteins encoded in the iscRSUA-hscBA-fdx-iscX-pepB-sseB, yadR/erpA and nfuA operons are significantly increased in this background, similar to that of the ΔiscR mutant strain (Figure S2A), indicating that holo-IscR represses expression of these genes under the aerobic, iron-replete conditions used. In contrast, increases in the sufABCDS Fe-S cluster biogenesis operon were observed for the apo-locked IscR strain when compared to both the wild type and ΔiscR strains (Figure S4). As IscR is overexpressed by 30-fold (p<0.05) in the apo-locked iscR mutant compared to wild type (Figure S2A), we speculate that the suf operon is positively regulated by IscR in Yersinia as in E. coli. In contrast, the extensively studied E. coli IscR target, hyaABCDEF, is not encoded in the Y. pseudotuberculosis genome.
Importantly, our RNAseq analysis demonstrated that transcription of genes within the virA, virB, virC, yscW-lcrF, and lcrGVH-yopBD operons was restored in the apo-locked IscR mutant compared to the ΔiscR mutant (Figure 7 and Table S2). However, we observed a decrease in transcription of genes encoding the T3SS effector proteins YopH (−4.4-fold), YopM (−3.0-fold), YopK (−7.1-fold), and YopE (−2.1-fold) in the apo-locked IscR mutant compared to wild type. Transcription of yopE has been shown to be regulated by Yop secretion through a positive feedback loop [51], [52], suggesting that the defect in YopHEMK transcription observed in the apo-locked IscR mutant may be caused by the lack of Yop secretion we observed in this strain. Together, these data suggest that both holo - and apo-IscR can promote T3SS gene transcription, possibly through binding to one or more type 2 DNA motifs.

To determine whether IscR might directly regulate T3SS gene expression, we carried out bioinformatic analysis to search pYV for sequences resembling the E. coli IscR type 2 motif (xxWWWWCCxYAxxxxxxxTRxGGWWWWxx) [30], [31], [33], as the DNA-binding domain of Yersinia IscR is 100% identical to that of E. coli IscR (Figure 1A). We searched within the 150 nucleotides upstream of the 99 genes encoded on the pYV plasmid and obtained a ranked list of putative type 2 motifs (data not shown). Among these was a site located within the yscW-lcrF promoter region (Figure 8A) [24]. To test whether IscR bound specifically to this site, we performed equilibrium DNA competition assays utilizing purified E. coli IscR-C92A (apo-locked IscR) [33], with a fluorescently-labeled E. coli hya type 2 site previously identified by Nesbit et al. [33]. Purified E. coli IscR was utilized in this assay, as complementation of the Y. pseudotuberculosis ΔiscR mutant strain with IscR of E. coli encoded on a plasmid fully restored secretion of T3SS cargo (Figure 8B). Competitor DNA included unlabeled E. coli hya as a positive control, the identified site within the Yersinia yscW-lcrF promoter region, a mutated version of this sequence (mlcrF), where nucleotides previously demonstrated in E. coli to be important for type 2 motif binding were altered [33], as well as one of the Y. pseudotuberculosis isc type 1 motif sites we identified as a negative control (Figure S2B & Figure 8C). We found that unlabeled lcrF DNA competed as well as unlabeled hya DNA (IC50 27 nm and 61 nm, respectively), suggesting that IscR can indeed bind to the identified type 2 motif upstream of lcrF (Figure 8D). Furthermore, mutation of key nucleotides in the lcrF promoter sequence led to alleviation of competition and increased the IC50 to greater than 1000 nM, a level comparable to that of the isc negative control type 1 motif site (Figure 8D). These findings suggest that IscR may regulate transcription of the T3SS through a type 2 motif within the yscW-lcrF promoter region.

Discussion
In this study, we present the first characterization of the iron-sulfur cluster regulator, IscR, of Yersinia. Initially identified through a genetic screen for modulators of Ysc T3SS function, iscR-deficient Y. pseudotuberculosis had a dramatic defect in secretion of T3SS effector proteins and in targeting macrophages through their T3SS, yet displayed normal growth in broth culture and wild type flagellar motility. Bioinformatic and DNA binding analysis revealed an IscR binding site upstream of the operon encoding the T3SS master regulator LcrF, indicating that IscR controls expression of the Ysc T3SS. Collectively, these findings indicated that IscR is a central component of the Y. pseudotuberculosis T3SS regulatory cascade.
Both E. coli holo - and apo-IscR are active transcription factors with distinct DNA binding targets. Holo-IscR can bind both type 1 and 2 motifs whereas apo-IscR can only bind type 2 motifs. IscR of E. coli autoregulates the isc operon, iscRSUA-hscBA-fdx, through binding to type 1 motifs within the isc promoter region [34]. In addition, Giel et al. described increased transcription of the genes located immediately downstream of the isc operon, yfhJ-pepB-sseB, in an iscR mutant, suggesting a negative regulatory effect on these genes as well [30]. We observed derepression of the iscRSUA-hscBA-fdx operon and the yfhJ-pepB-sseB locus in the Y. pseudotuberculosis ΔiscR mutant as well as the mutant expressing apo-locked IscR. Furthermore, we identified two sites within the Y. pseudotuberculosis isc promoter that closely match the E. coli IscR motif I consensus sequence. These data indicate that the iscRSUA-hscBA-fdx operon, and possibly the yfhJ-pepB-sseB locus, are negatively regulated by holo-IscR in Yersinia as they are in E. coli (Figure 9A). IscR in E. coli is known to activate transcription of the sufABCDSE operon through binding to a type 2 motif [29]. Our analysis revealed that the Y. pseudotuberculosis apo-locked IscR mutant overexpresses the sufABCDS operon compared to the wild type and ΔiscR strains, which we predict results from the overexpression of IscR observed in the apo-locked mutant as found in E. coli [32], [33]. We identified a site within the Y. pseudotuberculosis suf promoter region that closely resembles an E. coli IscR type 2 motif (data not shown). Together, these data indicate that the suf operon is positively regulated by IscR in Yersinia as in E. coli. Thus, we propose that IscR of Y. pseudotuberculosis modulates transcription of both the isc and suf Fe-S cluster biosynthesis pathways via mechanisms established for its E. coli ortholog.

In addition to control of Fe-S cluster biogenesis pathway expression, we present evidence that IscR controls expression and function of the Y. pseudotuberculosis T3SS. Bioinformatic analysis revealed a type 2 motif within the promoter of the T3SS master regulator LcrF that contained all nine bases previously found to be important for IscR binding (Figure 8A) [33]. Indeed, DNA binding assays demonstrated that IscR is able to specifically recognize this type 2 motif, suggesting that IscR may be acting directly to promote transcription of lcrF (Figure 9B). In support of this, we observed a marked decrease in transcription of numerous T3SS genes in the ΔiscR mutant strain. These include the gene that encodes LcrF, as well as a number of LcrF-regulated genes including the virC operon, yopK, yopT, yopM, yopH, yopJ, and lcrGVH-yopBD [17], [20], [22], [53], [54]. The lcrF type 2 motif is further upstream of the -10/-35 region previously identified by Böhme et al. [24] than other IscR binding sites that promote transcription [33], as we propose this site does. However, there may be an alternative −10/−35 region closer to the identified motif 2 site that might be used under specific growth conditions. Together, these data suggest that IscR is required for full expression of lcrF and LcrF-regulated genes through binding to a type 2 motif in the yscW-lcrF promoter (Figure 9B).
Based on these findings, an IscR mutant unable to coordinate an Fe-S cluster (apo-locked IscR) should lead to restoration of T3SS expression. Indeed, transcription of the yscW-lcrF and virC operons, as well as the majority of genes in the lcrGVH-yopBD operon, were no longer significantly decreased in the apo-locked IscR mutant compared to the ΔiscR strain. However, decreased transcription of yopE, yopK, yopM, and yopH as well as a severe defect in secretion of Yops was still observed. This could be explained by a deficiency in the apo-locked mutant's membrane potential, but not in the ΔiscR strain (Figure 9B). Wilharm et al., demonstrated that Y. enterocolitica motility and type III secretion requires the proton motive force [50]. Indeed, the apo-locked Y. pseudotuberculosis strain displayed a significant motility defect while the ΔiscR mutant was fully motile. Therefore, the type III secretion defect of the Y. pseudotuberculosis apo-locked IscR mutant can be explained by a deficiency in the proton motive force. Furthermore, the defect in YopHEMK transcription in the apo-locked IscR mutant may be explained by the fact that Yop secretion has a positive regulatory effect on Yop transcription [51], [52]. Together, these data suggest that apo-IscR can promote LcrF transcription, but that locking iscR is the apo form causes a proton motive force defect that prevents effector Yop transcription and secretion (Figure 9B).
It is unclear why locking IscR in the apo-locked form leads to a proton motive force defect. We observed ∼9-fold more suf transcript in the apo-locked IscR mutant compared to the ΔiscR strain that does not have a proton motive force defect, whereas the isc operon was expressed to the same degree in both mutants. Ezraty et al. recently showed that expression of the suf, but not the isc, operon in E. coli leads to a proton motive force defect, possibly as a result of impaired loading of Fe-S clusters into aerobic respiratory complexes [55]. Although the isc operon is expressed in the apo-locked Y. pseudotuberculosis mutant, perhaps overexpression of the suf pathway leads to misassembly of the Fe-S complexes of the electron transport chain that drive the proton motive force.
Both holo - and apo-IscR are predicted to bind to the type 2 motif within the yscW-lcrF promoter [33]. Based on previous data on E. coli IscR [28]–[30], [34], [56], low iron, aerobic growth, or high oxidative stress conditions are predicted to result in high expression of IscR through derepression of the isc operon, which in turn should increase T3SS gene expression. Likewise, high iron, anaerobic, or low oxidative stress conditions should lead to decreased IscR levels and therefore lower T3SS expression. Under normal aerobic culture conditions, we do not observe a change in wild type Y. pseudotuberculosis type III secretion when iron levels are altered (data not shown). However, in vivo, bacteria may be present in microaerophilic or anaerobic niches, where changes in iron bioavailability and reactive oxygen species production may impact iscR and T3SS gene expression. Upon ingestion by a host animal, Y. pseudotuberculosis enters the lumen of the intestine, which receives approximately 15 mg of iron per day [57], [58]. In the small intestine, Y. pseudotuberculosis can cross the gut barrier and enter the bloodstream and deeper tissues, which have very low iron bioavailability (∼10−24 M free serum iron) [59]–[61]. Sequestration of iron by iron carriers in mammalian tissues is an important host defense mechanism to prevent growth of bacterial pathogens, the majority of which require iron for growth [62]. The Ysc T3SS has been shown to be required for Y. pseudotuberculosis pathogenesis in these deep tissue sites that are low in iron bioavailability [44]. Perhaps Y. pseudotuberculosis uses IscR to sense iron, O2, and/or ROS concentration in order to optimally control T3SS expression in vivo.
Consistent with the severe T3SS expression defect displayed by the Y. pseudotuberculosis ΔiscR strain, this mutant was deficient in colonization of the Peyer's patches, spleen, and liver. Interestingly, the ΔiscR mutant was also defective in colonization of the mesenteric lymph nodes (MLN), yet T3SS mutants were previously shown to persist in the MLN and chromosomally-encoded factors were found to be important for Y. pseudotuberculosis survival in this tissue [24], [63], [64]. These results indicate that the virulence defect of the Y. pseudotuberculosis ΔiscR strain may not be due solely to misregulation of the T3SS, suggesting the existence of other IscR gene targets important for virulence. IscR of Pseudomonas aeruginosa has been shown to be important for full virulence through its ability to upregulate KatA, encoding a catalase that protects against oxidative stress [38], [65]–[67]. In Vibrio vulnificus, IscR upregulates two genes encoding the antioxidants peroxiredoxin and glutaredoxin 2, and is essential for survival during exposure to reactive oxygen species [40]. Interestingly, our analysis suggests that Y. pseudotuberculosis IscR plays an opposite regulatory role, as IscR negatively affects expression of the genes encoding cellular detoxification proteins KatY, Tpx, SodC and SodB. Furthermore, hydrogen peroxide sensitivity assays showed comparable levels of survival between the Y. pseudotuberculosis wild type and ΔiscR strains (Figure S5). This suggests that the virulence defect observed for the ΔiscR Y. pseudotuberculosis mutant is not due to increased susceptibility to oxidative stresses encountered during infection. Pathways other than the T3SS, such as the hmu hemin uptake system, were found to be misregulated in the Y. pseudotuberculosis ΔiscR strain (Table 2 & Figure 4B). While the hmu operon was shown to not affect Y. pestis virulence, it is possible that IscR control of the Y. pseudotuberculosis hmu pathway is important for virulence.
In summary, we present the first characterization for the iron-sulfur cluster regulator, IscR, of Yersinia. We reveal that IscR regulates genes involved in Fe-S cluster assembly in a manner akin to that of E. coli. Most notably, we demonstrate that mutation of IscR leads to decreased function of the Y. pseudotuberculosis T3SS and that this is due to a decrease in transcription of genes encoding structural, regulatory, and effector proteins. Furthermore, we present evidence showing that IscR is essential for the virulence of Y. pseudotuberculosis and that this attenuation is likely due, in part, to direct regulation of the T3SS by IscR. Collectively, this study argues for the important and novel role of IscR in the virulence of Y. pseudotuberculosis as well as regulation of the Ysc T3SS, and identifies IscR as a potential target for novel antimicrobial agents.
Materials and Methods
All animal use procedures were in strict accordance with the NIH Guide for the Care and Use of Laboratory Animals and were approved by the UCSC Institutional Animal Care and Use Committee.
Bacterial strains, plasmids and growth conditions
All strains used in this study are listed in Table 3. Y. pseudotuberculosis strains were grown in either 2xYT or M9 minimal media supplemented with casamino acids [68], referred to here as M9, at 26°C with shaking at 250 rpm, unless otherwise indicated. Where stated, Yop synthesis was induced via back-dilution of cultures into either M9 or low calcium media (2xYT plus 20 mM sodium oxalate and 20 mM MgCl2) to an OD600 of 0.2 and grown for 1.5 h at 26°C/shaking followed by 2 h at 37°C/shaking as previously described [69].

Construction of Y. pseudotuberculosis mutant strains
The iscR deletion mutant (ΔiscR) was generated via splicing by overlap extension PCR [70]. Primer pairs F5'iscR/R5'iscR and F3'iscR/R3'iscR (Table S3), designed using MacVector and Primer 3 software (http://fokker.wi.mit.edu/primer3/input.htm), were used to amplify ∼500 bp 5′ and 3′ of the iscR coding region, respectively. Amplified PCR fragments served as templates in an overlap extension PCR using the outside primers F5'iscR and R3'iscR.
The IscR-C92A/C98A/C104A mutant (apo-IscR) was generated via splicing by overlap extension PCR [70]. Primer pairs F5'apo-IscR/R5'apo-IscR and F3'apo-IscR/R3'apo-IscR (Table S3), designed using MacVector and Primer 3 software (http://fokker.wi.mit.edu/primer3/input.htm), were used to amplify ∼500 bp 5′ and 3′ within the iscR coding region, respectively. Amplified PCR fragments served as templates in an overlap extension PCR using the outside primers F5'apo-IscR and R3'apo-IscR. Nucleotide changes within the internal primers R5'apo-IscR and F3'apo-IscR allowed for amplification of iscR target containing sequences coding for alanine substitutions of the three conserved cysteines that coordinate an Fe-S cluster.
The resulting ∼1 kb fragments were cloned into the TOPO TA cloning vector (Invitrogen) and further subcloned into a BamHI - and NotI-digested pSR47s suicide plasmid (λpir-dependent replicon, kanamycinR (KanR), sacB gene conferring sucrose sensitivity) [71], [72]. Recombinant plasmids were transformed into E. coli S17-1 λpir competent cells and later introduced into Y. pseudotuberculosis IP2666 via conjugation. The resulting KanR, irgansanR (Yersinia selective antibiotic) integrants were grown in the absence of antibiotics and plated on sucrose-containing media to select for clones that had lost sacB (and by inference, the linked plasmid DNA). KanS, sucroseR, congo red-positive colonies were screened by PCR and subsequently sequenced to verify loss of the intended iscR coding region.
The iscR complement construct was generated by insertion of a fragment containing the iscR coding region as well as 530 bp of 5′ upstream sequence. This was PCR amplified using primer pair FiscRC and RiscRC, and cloned into the vector pACYC184 via BamHI/SalI restriction sites [73], [74]. Recombinant plasmids were transformed into E. coli S17-1 λpir competent cells and later introduced into Y. pseudotuberculosis IP2666 ΔiscR via a modified transformation method [75]. Briefly, recipient Yersinia strains were grown overnight in LB containing 2% glucose at 26°C. Cultures were centrifuged at 3,500 rpm for 3 min then washed with 750 µl of ice-cold sterile diH2O and repeated for a total of three washes. Washed pellets were resuspended in 100 µl of sterile diH2O, combined with 3 µl of plasmid and electroporated at EC2. Cells were allowed to recover in 1 mL SOC media for 1 h at 26°C followed by plating on LB containing carbenicillin to select for Yersinia bearing the plasmid of interest. Clones were confirmed by PCR analysis, using a combination of gene - and vector-specific primers, to construct both the ΔiscR complemented strain (ΔiscR pIscR) and the apo-IscR complemented strain (apo-IscR pIscR).
The nonmotile Δyop6/flhDCY.pestis mutant was generated by crossing in the Y. pestis flhDC gene into Y. pseudotuberculosis. Y. pestis flhD has a frameshift mutation, resulting in suppression of flagellin production [76]. The suicide plasmid pSB890 encoding a partial flhC gene and the full flhD gene from the Y. pestis KIM strain, generously provided by Dr. Brad Cookson, was conjugated into Y. pseudotuberculosis Δyop6 and nonmotile, recombinant mutants isolated as previously described [46].
Transposon screen generation and insertion site identification
Transposon mutagenesis was preformed similarly to Crimmins et al. [64]. Briefly, E. coli SM10λpir harboring pSC189, which encodes Himar1 [77], was mated with Y. pseudotuberculosis Δyop6. Mating culture was then pelleted, resuspended, spread out evenly among six 150 mm×15 mm petri plates containing LB supplemented with 2 µg mL−1 irgasan and 30 µg mL−1 Kan, and incubated for 3 days at room temperature. Colonies were patched onto LB supplemented with 100 µg mL−1 carbenicillin to ensure insertion of the transposon. Colony patches were used to grow 2xYT overnight cultures in 96-well plates, which were then frozen down to preserve the library. HEK293T cells were plated in 96-well white clear bottom plates (Corning) and transfected with a plasmid encoding a luciferase reporter gene fused to an NFκB-dependent promoter (Stratagene). Mutants from the transposon library were grown overnight in 96 well plates in M9 at 26°C and used to infect the transfected HEK293T cell monolayers. After 4 h incubation at 37°C, 100 µl of 1∶1 NeoLite:PBS solution was added to each well of the 96-well clear-bottom white plate (Corning), and luminescence was measured using a Victor3 plate reader (PerkinElmer). Each transposon mutant was assayed in duplicate. The positions of the transposons in the iscR::Tn1 and iscR::Tn2 mutants were determined by plasmid rescue, as previously described [78], except BamHI was used for digestion of genomic DNA.
NFκB activity assay
Validation of the transposon screen was performed through the use of an NFΚB activity assay, which is based on our previous work showing that Y. pseudotuberculosis induces NFκB activation in HEK293T cells dependent on expression of a functional T3SS [43]. Briefly, HEK293T cells were transfected with a plasmid encoding a luciferase reporter gene fused to an NFκB-dependent promoter (Stratagene). Bacterial strains were grown overnight in 2xYT and subcultured to an OD600 of 0.2 into low calcium media and grown at 26°C for 1.5 h followed by a shift to 37°C for an additional 1.5 h to induce the T3SS. Bacterial cultures were resuspended in prewarmed (37°C) DMEM and 200 µl aliquots were then used to infect the HEK293T cells containing the luciferase reporter plasmid at an MOI of 10. After 4 h incubation at 37°C, 100 µl of 1∶1 NeoLite:PBS solution was added to each well of the 96-well clear-bottom white plate (Corning), and luminescence was measured using a Victor3 plate reader (PerkinElmer). Data from three separate wells were averaged for each independent experiment.
Type III secretion assay
Visualization of T3SS cargo secreted in broth culture was performed as previously described [46]. Briefly, Y. pseudotuberculosis in M9 low calcium media (M9 plus 20 mM sodium oxalate and 20 mM MgCl2) was grown for 1.5 h at 26°C followed by growth at 37°C for 2 h. Cultures were normalized by OD600 and pelleted at 13,200 rpm for 10 min at room temperature. Supernatants were removed and proteins precipitated by addition of trichloroacetic acid (TCA) at a final concentration of 10%. Samples were incubated on ice for 20 min and pelleted at 13,200 rpm for 15 min at 4°C. Resulting pellets were washed twice with ice-cold 100% EtOH and subsequently resuspended in final sample buffer (FSB) containing 20% dithiothreitol (DTT). Samples were boiled for 5 min prior to running on a 12.5% SDS PAGE gel.
Ethidium bromide entry assay
Evaluation of pore formation was performed via the ethidium bromide (EtBr) entry assay as previously described [46]. Briefly, 2×104 immortalized C57Bl/6 BMDMs were plated in a 96 well clear bottom black plate (Corning) in 100 uL DMEM +10% FBS. Infection was performed in triplicate at an MOI of 25. Plates were centrifuged at 750×g at 4°C for 5 min to facilitate contact. Infections were carried out at 37°C with 5% CO2 for 2 h, at which point media was aspirated and replaced with 30 µL of PBS containing 25 µgmL−1 ethidium bromide (EtBr) and 12.3 µg mL−1 Hoechst dye. The cell monolayer was visualized using an ImageXpressMICRO automated microscope and MetaXpress analysis software (Molecular Devices). The percent of EtBr-positive cells was calculated by dividing the number of EtBr-stained cells by the number of Hoechst-stained cells. Data from three separate wells was averaged for each independent experiment.
Growth curves
Y. pseudotuberculosis strains were cultured overnight in 2xYT or M9 at 26°C and sub-cultured to an OD600 of 0.2 in 25 mL of either 2xYT or M9. Cultures were incubated at either 26°C or 37°C with shaking at 250 rpm and optical density measured at 600 nm every hour for 9 h.
Mouse infections
All animal use procedures were in strict accordance with the NIH Guide for the Care and Use of Laboratory Animals and were approved by the UC Santa Cruz Institutional Animal Care and Use Committee. Eleven to twelve-week-old 129S6/SvEvTac mice from our breeding facilities were used for oral infections as previously described [79]. Briefly, mice were orogastrically inoculated with 2×108 CFU in a 200 µl volume using a feeding needle. Mice were given food and water ad libitum and were euthanized at 5 days post-inoculation. Peyer's patches, mesenteric lymph nodes, spleens, and livers were isolated and homogenized for 30 s in PBS followed by serial dilution and plating on LB supplemented with 1 µg mL−1 irgasan for CFU determination.
RNAseq analysis
RNA was isolated from the IP2666 wild type and isogenic ΔiscR and apo-IscR strains grown for 3 h at 37°C in M9, using the RNeasy Mini Kit (Qiagen) as per the manufacturer's protocol. We chose M9 media for our RNASeq analysis because this condition enables expression of T3SS genes and secretion of T3SS cargo at 37°C [68]. Contaminating DNA was removed from the RNA samples using a DNA-free kit (Life Sciences). Samples were subjected to removal of contaminating rRNA via the Ribo-Zero Magnetic Kit for Gram-negative bacteria (Epicentre). The cDNA library was created using the NEBNext Ultra Directional RNA Library Prep Kit for Illumina (NEB). These studies were performed with three biological replicates per condition. Six indexed samples were sequenced per single lane using the HiSeq2500 Illumina sequencing platform for 50 bp single reads (UC Davis Genome Center) and subsequently analyzed and visualized via the CLC Genomics Workbench version 5.5.1 (CLC bio). Samples were normalized for both sequence depth and gene size by determining RPKM (Reads Per Kilo base per Million reads) and mapped to the Y. pseudotuberculosis genome (IP32953). Differentially regulated genes were identified as those displaying a fold change with an absolute value of 2 or greater. Statistical significance was determined by baySeq test with a corrected FDR post hoc test where p<0.05 was deemed significant [80].
Real-time PCR
Total RNA generated from our RNAseq analysis at a concentration of 2 µg was used to make cDNA as previously described [81]. SYBR Green PCR master mix (Applied Biosystems) was used for qPCR reactions according to the manufacturer's instructions and a 60°C annealing temperature. Primers used are listed in Table S4. Control primers were for the 16S rRNA as described previously [82]. Results were analyzed using the Bio-Rad CFX software.
Virulence plasmid map generation
Average RPKM values generated from RNAseq analysis for the wild type, ΔiscR and apo-IscR mutants were converted to log2-ratios (log2(RPKMmutant/RPKMwt) for each gene encoded on the virulence plasmid, pYV. These values were converted to a Circos heatmap [83] and plotted against the respective pYV base coordinate positions from Y. pseudotuberculosis IP32953.
Motif identification and search
Position specific scoring matrix (PSSM) was generated by the alignment of the known E. coli IscR type 2 motifs (Table S4) (Maverix Biomics, Inc) [31]. PSSM of type 2 was used to scan against the 150-nt upstream of 99 genes encoded on the Y. pseudotuberculosis pYV plasmid and obtained a ranked list of putative type 2 motifs.
DNA binding fluorescence anisotropy assays
Fluorescence anisotropy was measured similar to Nesbit et al., [33]. E. coli apo-IscR lacking the [2Fe-2S] cluster (IscR-C92A) was isolated anaerobically following the protocol described previously for wild type IscR [30]. Competition assays were performed using 5nM of 30-mer dsDNA of the known E. coli hyaA type 2 motif containing a 5′ TAMRA fluorophore (IDT) on the top strand and unlabeled competitor dsDNA concentrations ranged from 8 to 1000 nM (IDT, Table S4). DNA was annealed by heating equimolar concentrations of complementary DNA strands in annealing buffer (40 mM Tris (pH 7.9), 30 mM KCl) to 95°C for 5 min followed by slow cooling to room temperature over 2 hours. Annealed DNA was incubated with 90 nM apo-IscR in anisotropy buffer (40 mM Tris pH 7.9, 150 mM KCl, 100 ng ul−1 Salmon Sperm DNA) for 12 min at room temperature and anisotropy was measured using an EnVision 2103 Multilabel Reader (Perkin Elmer) with Wallac EnVision Manager software. Data is representative of experiments performed on three separate days.
Motility assay
Motility was analyzed by spotting 1 µl aliquots of either a nonmotile strain bearing an inactive, Y. pestis allele of flhD (Δyop6/flhDY.pestis), WT, ΔiscR, or apo-IscR strains onto motility agar plates (1% tryptone, 0.25% agar) from overnight cultures standardized to an OD600 of 2.5. Plates were incubated at room temperature for 1 day, at which point the diameters of the colonies were determined and used to calculate percent motility relative to WT, which was set at 100%.
Measurement of the membrane potential
The electrical potential was measured similar to the JC-1 red/green dye assay previously described for E. coli [84]. JC-1 is a membrane-permeable dye that emits green fluorescence (∼530 nm) upon excitation when the dye is in the monomeric form. Due to the membrane potential of the bacterial cell, JC-1 dye will form J aggregates which emit red fluorescence (∼590 nm). If the membrane potential decreases, there will be a decrease in J aggregate formation and subsequently a decrease in red fluorescence. As such, membrane potential can be displayed as a ratio of red/green fluorescence. Briefly, Y. pseudotuberculosis wild type and isogenic ΔiscR, ΔiscR complemented (ΔiscR pIscR), apo-IscR and apo-IscR complemented (apo-IscR pIscR) strains were grown overnight in M9 at 26°C. Strains were subcultured to an OD600 of 0.2 in M9 and grown at 37°C for 3 hours. A negative control containing a sample of wild type Y. pseudotuberculosis treated with 40 µM of the protonophore, CCCP (carbonyl cyanide m-chlorophenylhydrazone), during the last 30 min of growth was included in each experiment. After incubation at 37°C, 1 mL aliquots were harvested for each strain and pelleted at 4,500×g for 3 min followed by resuspension in 1 mL of permeabilisation buffer (10 mM Tris-HCl, pH 7.6, 1 mM EDTA and 10 mM glucose). Post resuspension, 2 µl of the membrane-permeable JC-1 dye (5 mg/mL) was added and the samples were incubated at room temperature for 30 min. Samples were then pelleted at 4,500×g for 3 min and resuspended in 500 µl of permeabilisation buffer. Slides were prepared by first coating with Poly-L-Lysine solution through addition of 100 µl aliquots of a 0.01% solution followed by a 5 min incubation at room temperature. Slides were washed a total of 3 times with sterile diH2O. Once dry, 10 µl of prepared sample was added to the slide and allowed to adhere for 5 min. Unattached bacteria were removed by washing with PBS and excess liquid removed via aspiration. A coverslip was applied and the cells were imaged using a LSM 5 PASCAL laser scanning microscope (Zeiss) fitted with a Plan-Apochromat 63x/1.4 Oil DIC objective and analyzed using the LSM 510 software (Zeiss). Quantification of image intensities was performed using ImageJ [85].
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. TroisfontainesP, CornelisGR (2005) Type III secretion: more systems than you think. Physiology (Bethesda) 20 : 326–339.
2. CoburnB, SekirovI, FinlayBB (2007) Type III secretion systems and disease. Clin Microbiol Rev 20 : 535–549.
3. ZhangY, MenaP, RomanovG, LinJS, SmileyST, et al. (2012) A protective epitope in type III effector YopE is a major CD8 T cell antigen during primary infection with Yersinia pseudotuberculosis. Infect Immun 80 : 206–214.
4. BliskaJB, WangX, ViboudGI, BrodskyIE (2013) Modulation of innate immune responses by Yersinia type III secretion system translocators and effectors. Cell Microbiol 15 : 1622–1631.
5. BrubakerRR (1983) The Vwa+ virulence factor of yersiniae: the molecular basis of the attendant nutritional requirement for Ca++. Rev Infect Dis 5 Suppl 4S748–758.
6. HueckCJ (1998) Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol Mol Biol Rev 62 : 379–433.
7. GemskiP, LazereJR, CaseyT, WohlhieterJA (1980) Presence of a virulence-associated plasmid in Yersinia pseudotuberculosis. Infect Immun 28 : 1044–1047.
8. RuckdeschelK, RoggenkampA, LafontV, MangeatP, HeesemannJ, et al. (1997) Interaction of Yersinia enterocolitica with macrophages leads to macrophage cell death through apoptosis. Infect Immun 65 : 4813–4821.
9. LarocheY, van BouchauteM, CornelisG (1984) A restriction map of virulence plasmid pVYE439-80 from a serogroup 9 Yersinia enterocolitica strain. Plasmid 12 : 67–70.
10. CornelisGR (2006) The type III secretion injectisome. Nat Rev Microbiol 4 : 811–825.
11. YipCK, StrynadkaNC (2006) New structural insights into the bacterial type III secretion system. Trends Biochem Sci 31 : 223–230.
12. DewoodyRS, MerrittPM, MarketonMM (2013) Regulation of the Yersinia type III secretion system: traffic control. Front Cell Infect Microbiol 3 : 4.
13. BlaylockB, RiordanKE, MissiakasDM, SchneewindO (2006) Characterization of the Yersinia enterocolitica type III secretion ATPase YscN and its regulator, YscL. J Bacteriol 188 : 3525–3534.
14. MuellerCA, BrozP, CornelisGR (2008) The type III secretion system tip complex and translocon. Mol Microbiol 68 : 1085–1095.
15. CornelisGR, BolandA, BoydAP, GeuijenC, IriarteM, et al. (1998) The virulence plasmid of Yersinia, an antihost genome. Microbiol Mol Biol Rev 62 : 1315–1352.
16. Lambert de RouvroitC, SluitersC, CornelisGR (1992) Role of the transcriptional activator, VirF, and temperature in the expression of the pYV plasmid genes of Yersinia enterocolitica. Mol Microbiol 6 : 395–409.
17. MichielsT, VanooteghemJC, Lambert de RouvroitC, ChinaB, GustinA, et al. (1991) Analysis of virC, an operon involved in the secretion of Yop proteins by Yersinia enterocolitica. J Bacteriol 173 : 4994–5009.
18. BergmanT, HakanssonS, ForsbergA, NorlanderL, MacellaroA, et al. (1991) Analysis of the V antigen lcrGVH-yopBD operon of Yersinia pseudotuberculosis: evidence for a regulatory role of LcrH and LcrV. J Bacteriol 173 : 1607–1616.
19. HoeNP, MinionFC, GoguenJD (1992) Temperature sensing in Yersinia pestis: regulation of yopE transcription by lcrF. J Bacteriol 174 : 4275–4286.
20. CornelisG, SluitersC, de RouvroitCL, MichielsT (1989) Homology between virF, the transcriptional activator of the Yersinia virulence regulon, and AraC, the Escherichia coli arabinose operon regulator. J Bacteriol 171 : 254–262.
21. CornelisG, VanootegemJC, SluitersC (1987) Transcription of the yop regulon from Y. enterocolitica requires trans acting pYV and chromosomal genes. Microb Pathog 2 : 367–379.
22. ChinaB, MichielsT, CornelisGR (1990) The pYV plasmid of Yersinia encodes a lipoprotein, YlpA, related to TraT. Mol Microbiol 4 : 1585–1593.
23. HoeNP, GoguenJD (1993) Temperature sensing in Yersinia pestis: translation of the LcrF activator protein is thermally regulated. J Bacteriol 175 : 7901–7909.
24. BohmeK, SteinmannR, KortmannJ, SeekircherS, HerovenAK, et al. (2012) Concerted actions of a thermo-labile regulator and a unique intergenic RNA thermosensor control Yersinia virulence. PLoS Pathog 8: e1002518.
25. RodionovDA, GelfandMS, ToddJD, CursonAR, JohnstonAW (2006) Computational reconstruction of iron - and manganese-responsive transcriptional networks in alpha-proteobacteria. PLoS Comput Biol 2: e163.
26. ShepardW, SoutourinaO, CourtoisE, EnglandP, HaouzA, et al. (2011) Insights into the Rrf2 repressor family—the structure of CymR, the global cysteine regulator of Bacillus subtilis. FEBS J 278 : 2689–2701.
27. FleischhackerAS, StubnaA, HsuehKL, GuoY, TeterSJ, et al. (2012) Characterization of the [2Fe–2S] cluster of Escherichia coli transcription factor IscR. Biochemistry 51 : 4453–4462.
28. WuY, OuttenFW (2009) IscR controls iron-dependent biofilm formation in Escherichia coli by regulating type I fimbria expression. J Bacteriol 191 : 1248–1257.
29. YeoWS, LeeJH, LeeKC, RoeJH (2006) IscR acts as an activator in response to oxidative stress for the suf operon encoding Fe-S assembly proteins. Mol Microbiol 61 : 206–218.
30. GielJL, RodionovD, LiuM, BlattnerFR, KileyPJ (2006) IscR-dependent gene expression links iron-sulphur cluster assembly to the control of O2-regulated genes in Escherichia coli. Mol Microbiol 60 : 1058–1075.
31. RajagopalanS, TeterSJ, ZwartPH, BrennanRG, PhillipsKJ, et al. (2013) Studies of IscR reveal a unique mechanism for metal-dependent regulation of DNA binding specificity. Nat Struct Mol Biol 20 : 740–747.
32. GielJL, NesbitAD, MettertEL, FleischhackerAS, WantaBT, et al. (2013) Regulation of iron-sulphur cluster homeostasis through transcriptional control of the Isc pathway by [2Fe-2S]-IscR in Escherichia coli. Mol Microbiol 87 : 478–492.
33. NesbitAD, GielJL, RoseJC, KileyPJ (2009) Sequence-specific binding to a subset of IscR-regulated promoters does not require IscR Fe-S cluster ligation. J Mol Biol 387 : 28–41.
34. SchwartzCJ, GielJL, PatschkowskiT, LutherC, RuzickaFJ, et al. (2001) IscR, an Fe-S cluster-containing transcription factor, represses expression of Escherichia coli genes encoding Fe-S cluster assembly proteins. Proc Natl Acad Sci U S A 98 : 14895–14900.
35. LeeKC, YeoWS, RoeJH (2008) Oxidant-responsive induction of the suf operon, encoding a Fe-S assembly system, through Fur and IscR in Escherichia coli. J Bacteriol 190 : 8244–8247.
36. BeinertH, HolmRH, MunckE (1997) Iron-sulfur clusters: nature's modular, multipurpose structures. Science 277 : 653–659.
37. Rincon-EnriquezG, CreteP, BarrasF, PyB (2008) Biogenesis of Fe/S proteins and pathogenicity: IscR plays a key role in allowing Erwinia chrysanthemi to adapt to hostile conditions. Mol Microbiol 67 : 1257–1273.
38. KimSH, LeeBY, LauGW, ChoYH (2009) IscR modulates catalase A (KatA) activity, peroxide resistance and full virulence of Pseudomonas aeruginosa PA14. J Microbiol Biotechnol 19 : 1520–1526.
39. Jones-CarsonJ, LaughlinJ, HamadMA, StewartAL, VoskuilMI, et al. (2008) Inactivation of [Fe-S] metalloproteins mediates nitric oxide-dependent killing of Burkholderia mallei. PLoS One 3: e1976.
40. LimJG, ChoiSH (2014) IscR is a global regulator essential for pathogenesis of Vibrio vulnificus and induced by host cells. Infect Immun 82 : 569–578.
41. MurphyER, PayneSM (2007) RyhB, an iron-responsive small RNA molecule, regulates Shigella dysenteriae virulence. Infect Immun 75 : 3470–3477.
42. EllermeierJR, SlauchJM (2008) Fur regulates expression of the Salmonella pathogenicity island 1 type III secretion system through HilD. J Bacteriol 190 : 476–486.
43. AuerbuchV, GolenbockDT, IsbergRR (2009) Innate immune recognition of Yersinia pseudotuberculosis type III secretion. PLoS Pathog 5: e1000686.
44. ViboudGI, BliskaJB (2005) Yersinia outer proteins: role in modulation of host cell signaling responses and pathogenesis. Annu Rev Microbiol 59 : 69–89.
45. KirbyJE, VogelJP, AndrewsHL, IsbergRR (1998) Evidence for pore-forming ability by Legionella pneumophila. Mol Microbiol 27 : 323–336.
46. KwuanL, AdamsW, AuerbuchV (2013) Impact of host membrane pore formation by the Yersinia pseudotuberculosis type III secretion system on the macrophage innate immune response. Infect Immun 81 : 905–914.
47. BliskaJB, GuanKL, DixonJE, FalkowS (1991) Tyrosine phosphate hydrolysis of host proteins by an essential Yersinia virulence determinant. Proc Natl Acad Sci U S A 88 : 1187–1191.
48. SheridanJJ, LogueCM, McDowellDA, BlairIS, HegartyT (1998) A study of the growth kinetics of Yersinia enterocolitica serotype O:3 in pure and meat culture systems. J Appl Microbiol 85 : 293–301.
49. GoverdeRL, KustersJG, Huis in 't VeldJH (1994) Growth rate and physiology of Yersinia enterocolitica; influence of temperature and presence of the virulence plasmid. J Appl Bacteriol 77 : 96–104.
50. WilharmG, LehmannV, KraussK, LehnertB, RichterS, et al. (2004) Yersinia enterocolitica type III secretion depends on the proton motive force but not on the flagellar motor components MotA and MotB. Infect Immun 72 : 4004–4009.
51. PetterssonJ, NordfelthR, DubininaE, BergmanT, GustafssonM, et al. (1996) Modulation of virulence factor expression by pathogen target cell contact. Science 273 : 1231–1233.
52. KauppiAM, NordfelthR, UvellH, Wolf-WatzH, ElofssonM (2003) Targeting bacterial virulence: inhibitors of type III secretion in Yersinia. Chem Biol 10 : 241–249.
53. WattiauP, CornelisGR (1994) Identification of DNA sequences recognized by VirF, the transcriptional activator of the Yersinia yop regulon. J Bacteriol 176 : 3878–3884.
54. SkurnikM, ToivanenP (1992) LcrF is the temperature-regulated activator of the yadA gene of Yersinia enterocolitica and Yersinia pseudotuberculosis. J Bacteriol 174 : 2047–2051.
55. EzratyB, VergnesA, BanzhafM, DuvergerY, HuguenotA, et al. (2013) Fe-S cluster biosynthesis controls uptake of aminoglycosides in a ROS-less death pathway. Science 340 : 1583–1587.
56. ZhengM, WangX, TempletonLJ, SmulskiDR, LaRossaRA, et al. (2001) DNA microarray-mediated transcriptional profiling of the Escherichia coli response to hydrogen peroxide. J Bacteriol 183 : 4562–4570.
57. McCanceRA, WiddowsonEM (1938) The absorption and excretion of iron following oral and intravenous administration. J Physiol 94 : 148–154.
58. MiretS, SimpsonRJ, McKieAT (2003) Physiology and molecular biology of dietary iron absorption. Annu Rev Nutr 23 : 283–301.
59. MartinRB, SavoryJ, BrownS, BertholfRL, WillsMR (1987) Transferrin binding of Al3+ and Fe3+. Clin Chem 33 : 405–407.
60. AisenP, LeibmanA, ZweierJ (1978) Stoichiometric and site characteristics of the binding of iron to human transferrin. J Biol Chem 253 : 1930–1937.
61. KretchmarSA, ReyesZE, RaymondKN (1988) The spectroelectrochemical determination of the reduction potential of diferric serum transferrin. Biochim Biophys Acta 956 : 85–94.
62. SkaarEP (2010) The battle for iron between bacterial pathogens and their vertebrate hosts. PLoS Pathog 6: e1000949.
63. Balada-LlasatJM, MecsasJ (2006) Yersinia has a tropism for B and T cell zones of lymph nodes that is independent of the type III secretion system. PLoS Pathog 2: e86.
64. CrimminsGT, MohammadiS, GreenER, BergmanMA, IsbergRR, et al. (2012) Identification of MrtAB, an ABC transporter specifically required for Yersinia pseudotuberculosis to colonize the mesenteric lymph nodes. PLoS Pathog 8: e1002828.
65. JangS, ImlayJA (2007) Micromolar intracellular hydrogen peroxide disrupts metabolism by damaging iron-sulfur enzymes. J Biol Chem 282 : 929–937.
66. JangS, ImlayJA (2010) Hydrogen peroxide inactivates the Escherichia coli Isc iron-sulphur assembly system, and OxyR induces the Suf system to compensate. Mol Microbiol 78 : 1448–1467.
67. FlintDH, TuminelloJF, EmptageMH (1993) The inactivation of Fe-S cluster containing hydro-lyases by superoxide. J Biol Chem 268 : 22369–22376.
68. ChengLW, AndersonDM, SchneewindO (1997) Two independent type III secretion mechanisms for YopE in Yersinia enterocolitica. Mol Microbiol 24 : 757–765.
69. MecsasJ, RaupachB, FalkowS (1998) The Yersinia Yops inhibit invasion of Listeria, Shigella and Edwardsiella but not Salmonella into epithelial cells. Mol Microbiol 28 : 1269–1281.
70. WarrensAN, JonesMD, LechlerRI (1997) Splicing by overlap extension by PCR using asymmetric amplification: an improved technique for the generation of hybrid proteins of immunological interest. Gene 186 : 29–35.
71. AndrewsHL, VogelJP, IsbergRR (1998) Identification of linked Legionella pneumophila genes essential for intracellular growth and evasion of the endocytic pathway. Infect Immun 66 : 950–958.
72. MerriamJJ, MathurR, Maxfield-BoumilR, IsbergRR (1997) Analysis of the Legionella pneumophila fliI gene: intracellular growth of a defined mutant defective for flagellum biosynthesis. Infect Immun 65 : 2497–2501.
73. ChangAC, CohenSN (1978) Construction and characterization of amplifiable multicopy DNA cloning vehicles derived from the P15A cryptic miniplasmid. J Bacteriol 134 : 1141–1156.
74. RoseRE (1988) The nucleotide sequence of pACYC184. Nucleic Acids Res 16 : 355.
75. WalkerKA, MillerVL (2004) Regulation of the Ysa type III secretion system of Yersinia enterocolitica by YsaE/SycB and YsrS/YsrR. J Bacteriol 186 : 4056–4066.
76. BergsbakenT, CooksonBT (2007) Macrophage activation redirects yersinia-infected host cell death from apoptosis to caspase-1-dependent pyroptosis. PLoS Pathog 3: e161.
77. ChiangSL, RubinEJ (2002) Construction of a mariner-based transposon for epitope-tagging and genomic targeting. Gene 296 : 179–185.
78. LawrenzMB, MillerVL (2007) Comparative analysis of the regulation of rovA from the pathogenic yersiniae. J Bacteriol 189 : 5963–5975.
79. AuerbuchV, IsbergRR (2007) Growth of Yersinia pseudotuberculosis in mice occurs independently of Toll-like receptor 2 expression and induction of interleukin-10. Infect Immun 75 : 3561–3570.
80. YaoJQ, YuF (2011) DEB: A web interface for RNA-seq digital gene expression analysis. Bioinformation 7 : 44–z45.
81. AuerbuchV, BrockstedtDG, Meyer-MorseN, O'RiordanM, PortnoyDA (2004) Mice lacking the type I interferon receptor are resistant to Listeria monocytogenes. J Exp Med 200 : 527–533.
82. ArafahS, RossoML, RehaumeL, HancockRE, SimonetM, et al. (2009) An iron-regulated LysR-type element mediates antimicrobial peptide resistance and virulence in Yersinia pseudotuberculosis. Microbiology 155 : 2168–2181.
83. KrzywinskiM, ScheinJ, BirolI, ConnorsJ, GascoyneR, et al. (2009) Circos: an information aesthetic for comparative genomics. Genome Res 19 : 1639–1645.
84. EnglC, BeekAT, BekkerM, de MattosJT, JovanovicG, et al. (2011) Dissipation of proton motive force is not sufficient to induce the phage shock protein response in Escherichia coli. Curr Microbiol 62 : 1374–1385.
85. SchneiderCA, RasbandWS, EliceiriKW (2012) NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9 : 671–675.
86. LarkinMA, BlackshieldsG, BrownNP, ChennaR, McGettiganPA, et al. (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23 : 2947–2948.
87. SchianoCA, BellowsLE, LathemWW (2010) The small RNA chaperone Hfq is required for the virulence of Yersinia pseudotuberculosis. Infect Immun 78 : 2034–2044.
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2014 Číslo 6
- Farmakovigilanční studie perorálních antivirotik indikovaných v léčbě COVID-19
- Jak souvisí postcovidový syndrom s poškozením mozku?
- Měli bychom postcovidový syndrom léčit antidepresivy?
- 10 bodů k očkování proti COVID-19: stanovisko České společnosti alergologie a klinické imunologie ČLS JEP
Nejčtenější v tomto čísle
- Fungal Nail Infections (Onychomycosis): A Never-Ending Story?
- Profilin Promotes Recruitment of Ly6C CCR2 Inflammatory Monocytes That Can Confer Resistance to Bacterial Infection
- IscR Is Essential for Type III Secretion and Virulence
- Contribution of Specific Residues of the β-Solenoid Fold to HET-s Prion Function, Amyloid Structure and Stability