
A Trans-Species Missense SNP in Is Associated with Sex Determination in the Tiger Pufferfish, (Fugu)
Heterogametic sex chromosomes have evolved independently in various lineages of vertebrates. Such sex chromosome pairs often contain nonrecombining regions, with one of the chromosomes harboring a master sex-determining (SD) gene. It is hypothesized that these sex chromosomes evolved from a pair of autosomes that diverged after acquiring the SD gene. By linkage and association mapping of the SD locus in fugu (Takifugu rubripes), we show that a SNP (C/G) in the anti-Müllerian hormone receptor type II (Amhr2) gene is the only polymorphism associated with phenotypic sex. This SNP changes an amino acid (His/Asp384) in the kinase domain. While females are homozygous (His/His384), males are heterozygous. Sex in fugu is most likely determined by a combination of the two alleles of Amhr2. Consistent with this model, the medaka hotei mutant carrying a substitution in the kinase domain of Amhr2 causes a female phenotype. The association of the Amhr2 SNP with phenotypic sex is conserved in two other species of Takifugu but not in Tetraodon. The fugu SD locus shows no sign of recombination suppression between X and Y chromosomes. Thus, fugu sex chromosomes represent an unusual example of proto–sex chromosomes. Such undifferentiated X-Y chromosomes may be more common in vertebrates than previously thought.
Published in the journal:
. PLoS Genet 8(7): e32767. doi:10.1371/journal.pgen.1002798
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002798
Summary
Heterogametic sex chromosomes have evolved independently in various lineages of vertebrates. Such sex chromosome pairs often contain nonrecombining regions, with one of the chromosomes harboring a master sex-determining (SD) gene. It is hypothesized that these sex chromosomes evolved from a pair of autosomes that diverged after acquiring the SD gene. By linkage and association mapping of the SD locus in fugu (Takifugu rubripes), we show that a SNP (C/G) in the anti-Müllerian hormone receptor type II (Amhr2) gene is the only polymorphism associated with phenotypic sex. This SNP changes an amino acid (His/Asp384) in the kinase domain. While females are homozygous (His/His384), males are heterozygous. Sex in fugu is most likely determined by a combination of the two alleles of Amhr2. Consistent with this model, the medaka hotei mutant carrying a substitution in the kinase domain of Amhr2 causes a female phenotype. The association of the Amhr2 SNP with phenotypic sex is conserved in two other species of Takifugu but not in Tetraodon. The fugu SD locus shows no sign of recombination suppression between X and Y chromosomes. Thus, fugu sex chromosomes represent an unusual example of proto–sex chromosomes. Such undifferentiated X-Y chromosomes may be more common in vertebrates than previously thought.
Introduction
Diverse systems of sex determination have evolved independently in the animal and plant kingdoms [1]. The most prominent of them involves heterogametic sex chromosome systems where whole or a part of sex chromosomes are heterozygous in one sex (XY or ZW) and homozygous in the other (XX or ZZ). In many species, these sex chromosomes show distinctive morphology. It has been hypothesized that these sex chromosomes originated from a pair of autosomes that eventually diverged due to suppression of recombination [2], [3]. In addition, a master sex-determining (SD) gene often resides on one of the sex chromosomes. However, the SD gene varies between organisms, which underscores the independent evolution of SD and sex chromosome systems [1]. In vertebrates, previously four master SD genes were known; Sry [4] in therian mammals; Dmrt1 in chicken [5]; Dmy, a duplicated copy of Dmrt1 in medaka [6]; and Dm-W, a truncated copy of Dmrt1 in Xenopus laevis [7]. All these SD genes code for transcription factors, belonging to either the Sox family (Sry) or the DM-domain family. When this paper was under review, two papers that identified non-transcription factors as vertebrate SD genes were published. One of these genes is a male-specific duplicated copy of the anti-Müllerian hormone (amhy) gene in the Patagonian pejerrey (Odontesthes hatcheri) [8]. The other is an allele of the gonadal soma derived growth factor (Gsdf) called GsdfY, located on the Y chromosome of Oryzias luzonensis, a species closely related to medaka [9]. Although studies of SD gene and sex chromosomes in vertebrates have provided significant insights into the evolution of sex-determining systems, our understanding is still very limited, given that sex chromosomes are known to exist in various states of differentiation [1] and that the SD genes remains to be identified in the vast majority of vertebrates.
In the present study, we investigated the SD region of the tiger pufferfish, Takifugu rubripes (fugu). Fugu is a large marine teleost and has an XX-XY sex determining system [10]. The availability of the whole genome sequence and a dense genetic map of fugu, combined with its compact genome size (400 Mb), allowed us to use the power of genetics to search for the SD gene in this wild species [11], [12].
Results/Discussion
Genetic Mapping
Our previous genome-wide linkage mapping in fugu has shown that the fugu SD region is restricted to a small segment of chromosome 19 flanked by large autosome-like regions [10], [12], [13]. In the fugu genome assembly (version 4), this locus includes four scaffolds of total length 5387 kb, containing ∼300 potential protein-coding genes [10]. Since a contiguous physical map of this region was not available, we first increased the resolution of the genetic map to determine the precise positions of the four scaffolds (Table S1). We next produced large numbers of fugu siblings and performed extensive linkage analyses to reduce the genomic interval of the SD region (Figure 1A, Table S2). We first narrowed down the SD region by analyzing 411 individuals and then searched for recombinants in the SD region in 1034 additional siblings. Comparison of the genotype of markers in the SD region and the sex of the 23 identified recombinants localized the SD region to a short stretch of 17.5 kb in which two protein-coding genes, NFX1-type zinc finger-containing 1 (Znfx1) and anti-Müllerian hormone receptor type II (Amhr2), are predicted (Figure 1B). Znfx1 is a transcription factor of unknown function that is expressed in a wide range of tissues in mammals including central nervous system, kidney, lung, muscle, ovary, testis, pancreas and thyroid [14], whereas Amhr2 is a receptor in the anti-Müllerian hormone (Amh) pathway that plays an important role in development and maintenance of reproductive organs in mammals and other vertebrates [15].
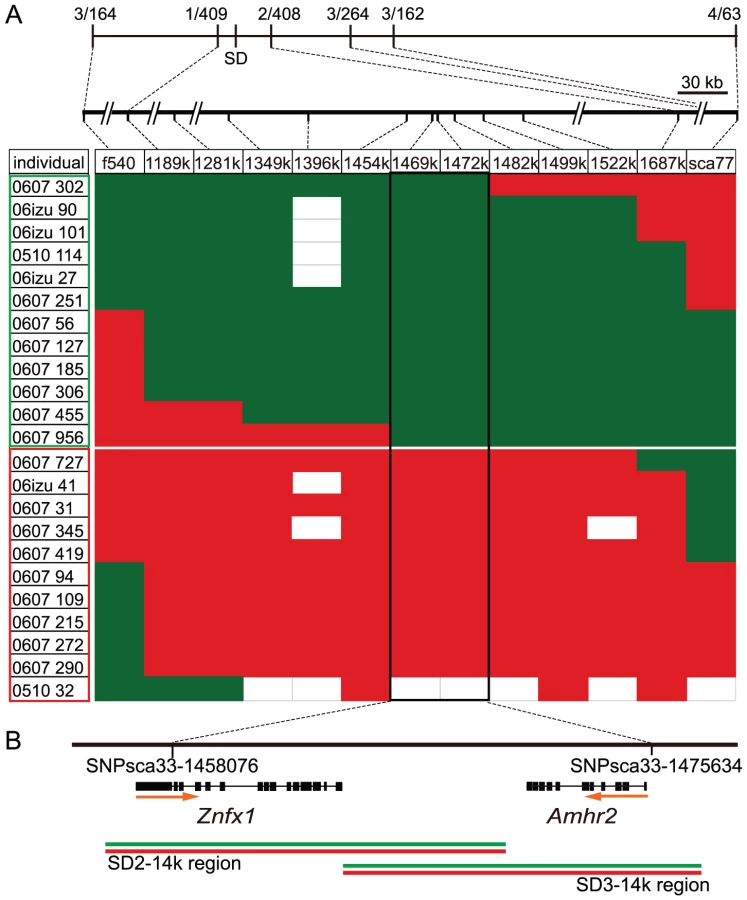
Since previously known SD genes in vertebrates are specific to one of the sex chromosomes (Y, Z or W) that shows varying degrees of sequence differentiation with the other sex chromosome [4]–[7], we examined whether the fugu SD region contains a distinct male-specific segment by PCR amplification, cloning and sequencing of the entire 17.5 kb SD region of two males chosen from experimental families (Figure 1B). We then determined whether the genomic clones were derived from X or Y by using the markers polymorphic in the families. The sequences of the X and Y derived clones covering the 17.5 kb SD region were more than 99% identical (99.4% for SD2–14k region and 99.6% for SD3–14K region, Figure S1) indicating that the male sex in fugu is determined by a small difference in the genomic sequence.
Association Mapping
To precisely pinpoint the SD gene, we employed association mapping utilizing ancestral recombination in a wild population of fugu (Figure 2). SNPs and other polymorphisms were screened by sequencing the entire SD region of seven males from seven experimental crosses, the 5′region of SD3 for X and Y in one male from an eighth family, and the 5′ region of SD3 for only X in a male from the ninth family (Figure S1, Table S3). In this analysis, we noted that only one of the variants, SNP 7271 (C>G), residing within exon 9 of Amhr2 is heterozygous in all males (Figure 3A, Table S3A and S3B). This SNP is located within the kinase domain of Amhr2 and changes an amino acid from His384 to Asp384 (Figure 3). Additional genotyping of this SNP in nine families showed perfect correlation with the male phenotype (Table S2). While 100% of males (41/41) were heterozygous (C and G) at this position, all females (44/44) were homozygous for the C allele.


Apart from this SNP, we did not find any other SNP or other types of variants (repeats, indels, etc.) that was heterozygous in any male (Table S3). We further investigated this SNP and other SNPs chosen from the 17.5 kb SD region and its neighboring regions (∼2 kb upstream harboring three SNPs and ∼600 bp downstream containing one SNP) in a natural population consisting of 58 females and 47 males (Figure 2). We first calculated linkage disequilibrium (LD) between all SNPs examined and found that the overall extent of LD in the SD region is very low (Figure 2C). This pattern suggested that recombination has not stopped in the region containing the fugu SD gene, unlike the previously characterized SD regions in other vertebrates [3]. Indeed, there is no significant LD even between markers that are 1 kb apart, thus providing a remarkably high resolution for the SD region. We then examined the association between each SNP and the sex phenotype. Consistent with the results of the small-scale analysis, SNP 7271 showed perfect association with the male phenotype (P = 5.6×10−31) (Figure 2B). In addition, its neighboring variant, SNP 7412 residing in intron 8 also showed a strong association with the male phenotype (P = 9.9×10−19). However, the latter is unlikely to be responsible for SD because 9 out of 105 wild fish showed discordance in their genotypes and phenotypic sex. The distribution of its genotype was 53 GG and 5 CG in females, and 4 GG, 40 CG, 2 CC and 1 undetermined in males. The association of SNP 7412 to phenotypic sex can be explained as an indirect association due to its close proximity to SNP 7271 (141 bp apart, r2 = 0.7) that shows a perfect correlation with the phenotypic sex (Figure 2 and Figure S2). The results of the association mapping studies together with those of variant screening suggest that the SNP 7271 (C>G) is the sole polymorphism in the 17.5 kb SD region that shows perfect correlation with phenotypic sex.
Trans-Species Comparison
Conservation of SNPs between species is rare, unless they are under some selective constraint, or that the species are so closely related that they still share their ancestor's variants. Thus, trans-species analysis offers an additional test to verify the correlation of SNP7271 to phenotypic sex in fugu [16], [17]. To this end, we investigated two other wild species of Takifugu, T. pardalis (n = 8 for each sex) and T. poecilonotus (n = 6 females and 7 males), that diverged from Takifugu rubripes approximately 5 million years ago (mya) [18]. We PCR amplified and sequenced a part of the Amhr2 gene (intron 8, exon 9 and intron 9) from them. Comparison of the sequences revealed that while SNP 7271 is present in both species, the intronic SNP 7412 is present only in T. pardalis (Figure 3B). More importantly, examination of all SNPs around exon 9 of Amhr2 from the two species indicated that SNP 7271 is the sole conserved polymorphism that shows perfect correlation with the sexual phenotype (Figure 3B and 3C and Figure S3, Table S4). These results strongly suggest that SNP 7271 is a trans-species SNP and is likely to be a causative variant for sex determination in the three species of Takifugu. Interestingly, this SNP is absent in the green spotted freshwater pufferfish, Tetraodon nigroviridis (n = 4 for each sex) which diverged from fugu approximately 40 to 70 mya [18],[19]. This indicates that the sex-determining polymorphism, SNP 7271 in Amhr2, evolved in a common ancestor of the three Takifugu species after it diverged from the Tetraodon lineage.
Structure and Identity of Fugu Amhr2 Gene
To confirm the exon-intron organization of the Amhr2 gene predicted in the fugu genome assembly, we amplified full-length cDNA of fugu Amhr2H384 and Amhr2D384 using cDNA from the testis as a template (accession number AB618627 in DDBJ). Sequencing of these products indicated that fugu Amhr2 comprises 11 exons, and encodes a protein of 514 amino acids (Figure S4A). The fugu protein is 28% identical to the human AMHR2 with 41% identity in the kinase domain (Figure S4A). The fugu Amhr2 gene is embedded in a syntenic block of genes that is conserved in Tetraodon, stickleback as well as in human (Figure S4B). This suggests that the fugu gene is an ortholog of Amhr2 in these species and that it was not transposed recently into the fugu SD locus in the Takifugu lineage. To further confirm its identity, we carried out phylogenetic analysis of Amhr2 and its related proteins from various vertebrates. This analysis confirmed that the fugu gene is an ortholog of Amhr2 in other vertebrates (Figure S4C). Searches of the draft genome assemblies of fugu, Tetraodon, stickleback and medaka identified only a single copy of Amhr2, suggesting that following the fish-specific whole genome duplication event in the teleost ancestor [20], [21], the duplicated copy of Amhr2 may have been lost in these teleosts.
Expression Pattern of Fugu Amhr2
In mammals, Amhr2 is responsible for the regression of the Müllerian duct in males [15]. Loss-of-function of this gene in male mouse leads to a partial hermaphrodite having a uterus and an oviduct together with the testis [15]. However, in medaka, a homozygous mutation in exon 9 of Amhr2 (Tyr390Cys) (hotei mutant) results in complete sex reversal in half of the genetic males [22]. These results indicate that although Amhr2 is not essential for formation of the testis in mammals, in fishes Amhr2 has the potential to influence the decision of the bipotential gonad to become either an ovary or a testis. If the Amhr2 gene indeed resides at the top of the sex determination pathway in fugu, it should be expressed before the differentiation of ovary and testis. However, it need not be expressed in a sex-specific manner like the mammalian Sry or medaka Dmy, because the sex-specific signaling can be produced by the male-specific isoform, Amhr2D384. Nevertheless, to verify if there is differential expression of Amhr2 between males and females, we analyzed Amhr2 expression in fugu by RT-PCR and in situ hybridization. Gonadal sex differentiation in fugu begins at 8–9 weeks after fertilization when juveniles attain a body length of approximately 25 mm [23]. The first sign of the morphological differentiation of ovary and testis is the formation of the ovarian cavity [23], [24]. The RT-PCR analysis revealed that Amhr2 is expressed in the differentiating ovary as well as the testis of juvenile fugu (Figure 4A; n = 3 for each sex). The in situ hybridization indicated that Amhr2 is expressed in both sexes before the morphological differentiation of gonads (Figure 4B; n = 3 for female and n = 4 for male), and subsequently in somatic cells surrounding the germ cells of the differentiating gonads (Figure 4B; n = 5 for each sex at 90 dpf and n = 2 for each sex at 126 dpf). A similar expression pattern of Amhr2 is also reported in medaka [25]. These results provide further support for a role for Amhr2 in the sex determination of fugu.

Effect of Missense SNP in Kinase Domain of Amhr2
Amhr2 is a type II receptor for the TGF-ß family of proteins and contains a single transmembrane domain and a serine/threonine kinase domain [26]. Upon binding to Amh, Amhr2 recruits and phosphorylates a type I receptor(s) that then transduces signals by phosphorylating Smad proteins which in turn regulate transcription of downstream genes in mammals [26]. SNP 7271 is located within the kinase domain of Amhr2 (Figure 3C) that is responsible for the phosphorylation. Five natural mutations in the kinase domain of Amhr2 in human [26] and one induced mutation in medaka (Tyr390Cys) [22] (see Figure S4D) resulted in a loss-of-function phenotype of Amh/Amhr2 signaling. Thus, the kinase domain is critical for the function of Amhr2. Since Asp384 is conserved in Tetraodon, stickleback, mouse and human (in medaka, Asp is replaced with Asn; Figure 3C), we propose that Amhr2H384 is a derived allele in Takifugu that causes a female phenotype when homozygous.
To determine the effect of Asp to His substitution in the kinase domain of Amhr2 on the activation of Smad proteins in fugu, we first tried an antibody that can recognize the activated Smad 1/5/8 in mammals and zebrafish on sections of the differentiating testis of fugu by immunohistochemistry [27]. However, the antibody was not effective in fugu. We then used a mouse teratocarcinoma cell line, P19, which contains all the molecules including type I receptor required for Amh signaling except Amhr2 [26] to assay the effect of substitution of fugu Amhr2 on the activation of Smad1. However, both fugu Amhr2D384 and Amhr2H384 failed to elicit Smad1 activity in this cell line (presumably due to the incompatibility with the mouse type I receptors) (data not shown). Because there are no comparable cell lines from fugu or other fishes, we introduced the His384 mutation into human AMHR2 (His397) and tested the human construct in P19 cells. Interestingly, the human mutant hAMHR2H397 mediated significantly less signaling (Smad1 activation) compared to hAMHR2D397 (Figure 5). This result suggests that fugu Amhr2H384 is an allele with reduced function compared to Amhr2D384 and the homozygous form of this less-effective allele facilitates the formation of ovary, whereas the heterozygous form of the alleles, Amhr2H384 and Amhr2D384, promotes development of testis. This implies that Amhr2D384 is a dominant allele. This interpretation is consistent with the male to female sex-reversal in the medaka Amhr2 homozygous mutant (hotei) that has a loss-of-function mutation in the kinase domain (Tyr390Cys) [22], located 13 positions downstream of fugu Asp384 (see Figure 3C).

One possible mechanism of sex determination by Amh/Amhr2 signaling is that it decreases the number of germ cells, which in turn promotes the development of testis. This hypothesis is based on the observation that in the medaka hotei mutant over-proliferated germ cells were associated with ovary formation in males [22]. On the other hand, inhibition of primordial germ cell migration into gonads by knockdown of the chemoattractant receptor gene, cxcr4, resulted in the formation of testis-like structure in females [28]. Consistent with this finding, high temperature-induced germ cell degeneration in fugu is associated with masculinization of ovarian somatic cells [29]. Another possibility is that the Amh/Amhr2 signaling could antagonistically act on the aromatase activity as reported in mammals [30]. Enhanced estrogen synthesis by aromatase is known to function as a natural inducer of ovarian differentiation in fishes [31]. Thus suppression of aromatase activity by Amh/Amhr2 signaling could lead to the formation of testis in fugu.
Allelic Sex Determination by a Non-Transcription Factor
In vertebrates, previously four master SD genes were known: Sry in therian mammals [4]; Dmrt1 in chicken [5]; Dmy in medaka [6]; and Dm-W in Xenopus laevis [7]. They all code for transcription factors and are able to regulate the expression of other genes in the sex-determination pathway. By contrast, the Amhr2 gene associated with the sex determination in fugu codes for a growth factor receptor. This suggests that any gene that lies in the sex-determination pathway has the potential to be recruited for triggering sex determination. Furthermore, unlike the known vertebrate SD genes that reside on only one of the pair of sex chromosomes, fugu Amhr2 is located on both of the homologous chromosomes. This suggests that phenotypic sex in fugu is determined by a combination of the two allelic variants of Amhr2 gene.
When this paper was under review, two other novel candidates for vertebrate SD genes were reported, and both are in fishes. The first one is a male-specific, duplicated copy of the Amh gene (amhy) implicated in testicular development of the Patagonian pejerrey [8]. Morpholino-mediated knock down of amhy (and presumably amh) resulted in male to female sex reversal in 22% of fishes carrying the duplicated copy of Amh. Interestingly, Amh codes for a hormone that is a ligand for Amhr2. The other SD gene, GsdfY identified in O. luzonensis [9], is an allele similar to the sex-determining Amrh2 allele in fugu. The promoter analysis of this gene showed that the allelic difference involves 6 to 9 nucleotide difference in the binding site for a steroidogenic factor 1 (SF1) and results in male-specific high expression of the GsdfY allele. By contrast, the polymorphism in fugu is in the coding sequence and affects the function of the protein. Nevertheless, it is interesting that all these three novel fish sex-determining genes are components of the TGF-ß signaling pathway. These findings underscore the critical role of TGF-ß signaling, a hitherto unappreciated pathway, in gonadal sex determination in vertebrates. It is possible that additional members of this pathway may be involved in sex determination in other teleosts and other non-mammalian vertebrates that have experienced a recent turnover of sex chromosomes.
Establishment of a Recessive Allele as a New SD Gene
Several models have been proposed to explain the evolution of new sex-determination mechanisms. These include random genetic drift [32], pleiotropic selection favoring new SD alleles [33] and sexually antagonistic selection [34]. The last model begins with an autosomal locus segregating alleles that have different relative fitness in males and females. Selection will favor an increase in the frequency of any new SD locus that is linked to these sexually antagonistic alleles. When the new SD allele is recessive, like the fugu Amhr2H384, it cannot increase in frequency quickly. However, a recessive allele could first increase in frequency by drift. Once its frequency reaches a certain level, sexually antagonistic selection at the sex-linked locus can lead to the recessive feminizing allele to spread to fixation, resulting in the disappearance of the previous female determiner such as a dominant allele on the W chromosome. This could lead to heterogamety switch from ZW/ZZ to XX/XY. Since Amh/Amhr2 signaling plays a role downstream of the SD gene in many vertebrates including mammals, birds and medaka [15], [22], [35], the fixation of such a downstream factor as the new SD locus might explain the later onset of the gonadal dimorphism, which occurs around 8–9 weeks after fertilization in fugu.
Proto–Sex Chromosomes
The theory of sex chromosome predicts that when an existing SD gene is replaced by a new SD gene, the young sex chromosomes lack a nonrecombining region. Such a region subsequently evolves through suppression of recombination leading to the divergence of the sequences of the homologous sex chromosomes [3]. The suppression of recombination is generally mediated by accumulation of repetitive sequences and/or inversion of chromosomal segments spanning the newly arisen SD gene/locus [36]. The sex chromosome theory thus implies that just after a new SD gene has replaced an existing one, the two homologous chromosomes show little divergence in their sequences even around the SD gene. However, no such instance of highly similar sex chromosome pair is known in vertebrates except in fugu, and the recently reported O. luzonensis [9]. The two youngest known SD genes in vertebrates, medaka Dmy and Xenopus Dm-W, arose by segmental duplications and are embedded within recognizable nonrecombining regions. In medaka, the Y-specific nonrecombining region, originally derived from the duplication of a 43-kb fragment of the Dmrt1 locus, has grown to 258 kb by accumulating 137 kb of repetitive sequences [37]. The W-specific region of Xenopus, likely derived from a duplication of the Dmrt1 locus, is flanked by 3 to 4 kb of nonrecombining regions [7], [38].
In fugu, although the recombination ratio in males is reduced compared to females even in regions around SD locus (Table S1), our genetic analysis did not identify any nonrecombining region in the SD locus (Figure 1 and Figure 2). This is consistent with our previous observation that the suppression of recombination in males relative to females is no greater around the sex-determination locus than on autosomes [12]. Furthermore, sequence comparison between the two SD chromosomes did not reveal any inversions or large scale accumulation of transposons and other repetitive elements (Figure 1 and Figure S1). Overall, the fugu SD region contains very little repetitive sequences (1.8% of retroelements and 2.3% simple repeats). Therefore, the SD chromosomes of fugu provide an unusual example of proto-sex chromosomes. The SD locus in therian mammals, medaka and Xenopus laevis are associated with duplication events, which can explain recombination suppression. However, in the case of Takifugu species, since sex is determined by just a single nucleotide difference, the SD locus might have eluded recombination suppression. Although there is little information regarding recombination in the SD locus of O. luzonensis [9], it would be interesting to determine if recombination around this SD gene still occurs, as in fugu.
Stock et al. [39] have recently shown that three species of European tree frogs that diverged 5 to 7 mya share the same pair of sex chromosomes with complete absence of X-Y recombination in males. Yet, the sequences of sex-linked loci are very similar between the X and Y chromosomes. Since phylogenetic analysis showed that the X and Y alleles clustered according to species rather than gametologs, the authors proposed that the similarity of the sequences at the sex-linked loci is maintained by occasional X-Y recombination, presumably occurring in sex-reversed XY females. Although this model may explain the sequence similarity between X and Y at the SD locus in fugu, the frequency of XY fugu females appears to be very low. We have not encountered any XY female in our genetic experiments. Moreover, attempts at producing XY females by hormonal or temperature manipulation have proven to be unsuccessful [29], [40]. Thus, the extent of the contribution of sex-reversed XY females in maintaining similarity at the SD locus of fugu is unclear.
Conclusion
We investigated the SD locus in fugu by high-resolution genetic mapping and association mapping. We found that a missense SNP in the kinase domain of Amhr2 that changes an amino acid is the sole polymorphism perfectly correlated with phenotypic sex. Our results suggest that a combination of the two alleles of Amhr2 is responsible for sex determination in fugu. The pattern of LD across the fugu SD locus indicates the absence of a nonrecombining region. Thus, the sex chromosomes of fugu represent a unique example of proto-sex chromosomes in vertebrates. Since genetic mapping studies of sex-determination loci in diverse taxa of vertebrates have indicated that vertebrates such as fishes, reptiles and amphibians have experienced a rapid turnover of sex chromosomes [1], the pre-differentiated sex chromosomes found in fugu may be more common among these vertebrates than previously thought.
The successful identification of the candidate sex-determining SNP in this study relied essentially on a sub-gene-level resolution of the association mapping owing to the low degree of LD in a wild population like fugu. In human populations, this method usually provides resolution at the level of only one or a few genes (tens to hundreds of kb) [41]. The resolution is even lower in laboratory mouse and domestic animals due to the strong LD across the genomes caused by a small effective population size [42]. By contrast, a wild species with a large effective population size is likely to show a low degree of LD across its genome. Thus the use of association mapping should greatly facilitate the identification of SD genes and chromosomes in the wild populations of non-model vertebrates.
Materials and Methods
Pedigreed Families for Linkage Mapping
To produce siblings, we crossed a fugu male with a fugu female as described previously [13]. Details of the families used in linkage mapping are shown in Table S2. Sex was determined by histological examination of the gonads for fishes aged 2–10 months and by visual inspection for older fish. The microsatellite loci were chosen by scanning scaffolds as described previously [13]. Primer sequences for microsatellite markers are listed in Table S5. Genotyping with microsatellite markers was performed as described previously [13]. This project was conducted in accordance with the Regulation for Animal Experiments of the University of Tokyo.
DNA Sequences of the SD Region from X and Y Chromosomes
We generated nine families and used males from these pedigrees for generating complete sequences of the SD region from X and Y chromosomes (Figure S1, Table S2). Genomic clones covering Znfx1 (genomic region of SD2–14k) and Amhr2 (genomic region of SD3–14k) were obtained from the XY single male by PCR using KOD FX (Toyobo) reagents suitable for amplifying large genomic fragments (Figure 1B and Figure S1). Approximately 50 ng of genomic DNA was used as a template in 25 µl PCR reaction. PCR amplification was performed using KOD DNA polymerase (Toyobo) with the primer pair SD2–14.7kF and SD2–14.7kR for SD2–14k genomic region, and the primer pair 33–1464k340F and 33–1464k13469R for SD3–14k genomic region. The cycling conditions were 36 cycles of 94°C for 10 s and 70°C for 12 min. Primer sequences are given in Table S5. DNA fragments were cloned by using TOPO XL PCR cloning kit (Invitrogen) or In-Fusion Advantage PCR cloning Kit (Clontech). The clones from X and Y chromosome were distinguished by SNP7271 or satellite marker 1469 K and were subjected to sequencing. To identify candidate polymorphic sites and obtain variants for linkage disequilibrium analysis, we cloned genomic DNA covering Amhr2 gene (the 5′ and 3′ regions of SD3, Figure S1) from males from seven independent families. In addition, we cloned genomic DNA covering the 5′ region of SD3 from two males from two other independent families. We sequenced theses clones and identified variants among them (Table S3). To avoid cloning artifacts, we followed the method described by Saitoh and Chen [43]. For variants found in only one clone, two additional clones were sequenced to obtain the consensus sequence. For the region covering Znfx1 (genomic region of SD2–14k), we directly sequenced the PCR products from ten males and determined DNA sequences for at least seven individuals from ten males.
Association Test
We studied a natural population of fugu consisting of 58 females and 47 males from off shore areas around the mid-west part of Japan. Genotyping was done using TaqMan or restriction fragment length polymorphism (RFLP) analysis (Table S4). We calculated association for SNPs having a minor allele frequency (MAF)>0.1 and a call rate >99% with phenotypic sex in fugu using Haploview program 4.1 [44]. Uncorrected P-values and P-values with 1,000,000 permutations are reported. We also tested the association between genotypes and phenotypic sexes for a recessive model of penetrance in which homozygosity of one of the alleles is required for phenotypic female based on the previous studies that have suggested that the sex of fugu is determined by an XX-XY system [10]. Linkage disequilibrium plot of r2 and D′ was generated using Haploview.
cDNA Cloning and Phylogenetic Analysis
cDNA of the fugu Amhr2H384 and Amhr2D384 were cloned from the testis using SMART RACE cDNA amplification Kit (Clontech) and sequenced completely. Multiple sequence alignment and NJ tree of fugu Amhr2 and its related proteins from various vertebrates were generated using ClustalW [45].
Trans-Species Association Test
We obtained T. pardalis (n = 8 for each sex) and T. poecilonotus (6 females and 7 males) from Lake Hamana, Japan. To determine exon 9 sequence of Amhr2 and its neighboring regions, we first amplified the genomic region using primer SD3exon8F and SD3exon10R, and sequenced the PCR product directly. Tetraodon nigroviridis specimens (n = 4 for each sex) were obtained from a supplier in Philippines, and the sequence of Amhr2 exon 9 and its neighboring regions were determined after PCR amplification with primers Tet-SD3exon8F and Tet-SD3exon10R. See Table S5 for primer sequences.
RT–PCR
One microgram of total RNA each from the gonad, brain, eye, intestine, heart, liver, spleen, kidney, trunk muscle and skin of fugu at 147 days post-fertilization (dpf) was used for synthesizing first-strand cDNA. PCR amplification was performed with the primer pair RT-SD3exon9F and RT-SD3exon9R, and the primer pair EF1α-F and EF1α-R. The cycling conditions were 40 cycles of 94°C for 10 s, 58°C for 5 s, and 72°C for 30 s. See Table S5 for primer sequences.
In Situ Hybridization
Juveniles at 62, 90 and 126 dpf from a full-sib family were dissected along the ventral midline and fixed in Bouin's solution or 4% paraformaldehyde at 4°C overnight. Fixed samples were dehydrated in graded ethanol, embedded in paraffin and sectioned serially at 5 µm thickness. After rehydration, sections were subjected to hybridization. The probes were transcribed from a fugu Amhr2 cDNA construct using DIG RNA labeling kit (Roche). Signals were detected by immunoreaction with alkaline phosphatase-conjugated anti-DIG antibody and NBT/BCIP solution (Roche). The genotypic sexes of all fish were determined by SNP7271 while phenotypic sex of fish at 90 and 126 dpf were determined by the formation of the ovarian cavity. The ovarian cavity was not seen in any fish at 64 dpf with the body length ranging from 19 to 22 mm (n = 4 for XY fish and n = 5 for XX fish).
Constructs for Luciferase Assay
The coding region of human AMHR2 was PCR amplified using a cDNA clone (MHS4426-99239518, Open Biosystems) as a template with hAMHRII-F and hAMHRII-R primers. The amplicon was ligated into the pcDNA3.1 vector (Invitrogen). Site-directed mutation was carried out using KOD-Plus-Mutagenesis Kit (Toyobo) with hAMHRII-D397H-F and hAMHRII-D397H-R primers. The coding region of mutated hAMHRII was amplified by PCR and ligated into the pcDNA3.1. Coding region of human Smad1 was amplified by PCR using Smad1f and Smad1r primers with pCMV5 Smad1 (Addgene) as a template. The amplicon and the pBIND vector (Promega) were ligated to generate the construct Gal4-Smad1/pBIND. The constructs were confirmed by sequencing. Construction of 5xGal4-tk-luc construct has been reported previously [46]. See Table S5 for primer sequences.
Cell Culture of P19 and Luciferase Assay
Mouse embryonal carcinoma P19 cells were cultured in Minimum Essential Medium (αMEM, Sigma M8042) containing 2 mM L-Alanyl-L-Glutamine, 7.5% FCS and 2.5% FBS at 37°C under 5% CO2. P19 was seeded in 96-well plates at 1×104 cells/100 µl medium 24 h prior to transfection. Transfection was conducted using Lipofectamine LTX (Invitrogen). 5xGal4-tk-luc, Gal4-Smad1/pBIND and empty pcDNA3.1, wild-type or mutated human AMHR2 in pcDNA3.1 were transfected. After addition of the recombinant human MIS (rhMIS, R&D Systems, 2 µg/ml), the cells were cultured for 24 h. Luciferase activity was determined using Dual Luciferase Kit (Promega). Firefly luciferase activity was normalized to Renilla luciferase activity. Each experiment was done in triplicates and the average of normalized activity was calculated. Three independent transfections were carried out, and values obtained by rhMIS treatment were divided by those obtained without rhMIS treatment. Data were expressed as mean ± SEM, with mean values comprising values from three independent assays: 3 per assay (9 per treatment). Luciferase activity was compared by One-way ANOVA (two-tailed, P<0.05). The Holm-Sidak test was applied to measure the differences among means for those cell types (overall significance level P = 0.05).
Supporting Information
{kind=link}
{kind=link}
Zdroje
1. CharlesworthDMankJE 2010 The birds and the bees and the flowers and the trees: lessons from genetic mapping of sex determination in plants and animals. Genetics 186 9 31
2. OhnoS 1967 Sex chromosomes and sex-linked genes Berlin, New York etc. Springer-Verlag 192
3. BergeroRCharlesworthD 2009 The evolution of restricted recombination in sex chromosomes. Trends Ecol Evol 24 94 102
4. SinclairAHBertaPPalmerMSHawkinsJRGriffithsBL 1990 A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 346 240 244
5. SmithCARoeszlerKNOhnesorgTCumminsDMFarliePG 2009 The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature 461 267 271
6. MatsudaMNagahamaYShinomiyaASatoTMatsudaC 2002 DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 417 559 563
7. YoshimotoSOkadaEUmemotoHTamuraKUnoY 2008 A W-linked DM-domain gene, DM-W, participates in primary ovary development in Xenopus laevis. Proc Natl Acad Sci USA 105 2469 2474
8. HattoriRSMuraiYOuraMMasudaSMajhiSK 2012 A Y-linked anti-Müllerian hormone duplication takes over a critical role in sex determination. Proc Natl Acad Sci USA 109 2955 2959
9. MyoshoTOtakeHMasuyamaHMatsudaMKurokiY 2012 Tracing the emergence of a novel sex-determining gene in medaka, Oryzias luzonensis. Genetics doi:10.1534/genetics.111.137497
10. KikuchiKKaiWHosokawaAMizunoNSuetakeH 2007 The sex-determining locus in the tiger pufferfish, Takifugu rubripes. Genetics 175 2039 2042
11. AparicioSChapmanJStupkaEPutnamNChiaJM 2002 Whole-genome shotgun assembly and analysis of the genome of Fugu rubripes. Science 297 1301 1310
12. KaiWKikuchiKTohariSChewAKTayA 2011 Integration of the genetic map and genome assembly of fugu facilitates insights into distinct features of genome evolution in teleosts and mammals. Genome Biol Evol 3 424 442
13. KaiWKikuchiKFujitaMSuetakeHFujiwaraA 2005 A genetic linkage map for the tiger pufferfish, Takifugu rubripes. Genetics 171 227 238
14. YanaiIBenjaminHShmoishMChalifa-CaspiVShklarM 2005 Genome-wide midrange transcription profiles reveal expression level relationships in human tissue specification. Bioinformatics 21 650 659
15. JaminSPArangoNAMishinaYHanksMCBehringerRR 2003 Genetic studies of the AMH/MIS signaling pathway for Müllerian duct regression. Mol Cell Endocrinol 211 15 19
16. CharlesworthD 2006 Balancing selection and its effects on sequences in nearby genome regions. PLoS Genet 2 e64 doi:10.1371/journal.pgen.0020064
17. WiufCZhaoKInnanHNordborgM 2004 The probability and chromosomal extent of trans-specific polymorphism. Genetics 168 2363 2372
18. YamanoueYMiyaMMatsuuraKMiyazawaSTsukamotoN 2009 Explosive speciation of Takifugu: another use of fugu as a model system for evolutionary biology. Mol Biol Evol 26 623 629
19. BentonMJDonoghuePC 2007 Paleontological evidence to date the tree of life. Mol Biol Evol 24 26 53
20. HoeggSBrinkmannHTaylorJSMeyerA 2004 Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J Mol Evol 59 190 203
21. ChristoffelsAKohEGLChiaJBrennerSAparicioS 2004 Fugu genome analysis provides evidence for a whole-genome duplication early during the evolution of ray-finned fishes. Mol Biol Evol 21 1146 1151
22. MorinagaCSaitoDNakamuraSSasakiTAsakawaS 2007 The hotei mutation of medaka in the anti-Müllerian hormone receptor causes the dysregulation of germ cell and sexual development. Proc Natl Acad Sci USA 104 9691 9696
23. MatsuuraSNaitoTShinchoMYoshimuraKMatsuuraM 1994 Gonadal sex differentiation in tiger puffer, Takifugu rubripes. SUISANZOSHOKU 42 619 625
24. YamaguchiALeeKHFujimotoHKadomuraKYasumotoS 2006 Expression of the DMRT gene and its roles in early gonadal development of the Japanese pufferfish Takifugu rubripes. Comp Biochem Physiol D 1 59 68
25. ShiraishiEYoshinagaNMiuraTYokoiHWakamatsuY 2008 Müllerian inhibiting substance is required for germ cell proliferation during early gonadal differentiation in medaka (Oryzias latipes). Endocrinology 149 1813 1819
26. BelvilleCMaréchaJDPennetierSCarmilloPMasgrauL 2009 Natural mutations of the anti-Müllerian hormone type II receptor found in persistent Müllerian duct syndrome affect ligand binding, signal transduction and cellular transport. Hum Mol Genet 18 3002 3013
27. NeumannJCChandlerGLDamoulisVAFustinoNJLillardK 2011 Mutation in the type IB bone morphogenetic protein receptor Alk6b impairs germ-cell differentiation and causes germ-cell tumors in zebrafish. Proc Natl Acad Sci USA 108 13153 13158
28. KurokawaHSaitoDNakamuraSKatoh-FukuiYOhtaK 2007 Germ cells are essential for sexual dimorphism in the medaka gonad. Proc Natl Acad Sci USA 104 16958 16963
29. LeeKYamaguchiARashidHKadomuraKYasumotoS 2009 Germ cell degeneration in high-temperature treated pufferfish, Takifugu rubripes. Sex Dev 3 225 232
30. di ClementeNGhaffariSPepinskyRPieauCJossoN 1992 A quantitative and interspecific test for biological activity of anti-Müllerian hormone: the fetal ovary aromatase assay. Development 114 721 727
31. DevlinRHNagahamaY 2002 Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental influences. Aquaculture 208 191 364
32. VuilleumierSLandeRJJVANASeehausenO 2007 Invasion and fixation of sex-reversal genes. J Evol Biol 20 913 920
33. PomiankowskiANothigerRWilkinsA 2004 The evolution of the Drosophila sex-determination pathway. Genetics 166 1761 1773
34. van DoornGSKirkpatrickM 2007 Turnover of sex chromosomes induced by sexual conflict. Nature 449 909 912
35. SmithCASinclairAH 2004 Sex determination: insights from the chicken. Bioessays 26 120 132
36. FraserJAHeitmanJ 2005 Chromosomal sex-determining regions in animals, plants and fungi. Curr Opin Genet Dev 15 645 651
37. KondoMHornungUNandaIImaiSSasakiT 2006 Genomic organization of the sex-determining and adjacent regions of the sex chromosomes of medaka. Genome Res 16 815 826
38. BewickAJAndersonDWEvansBJ 2011 Evolution of the closely related, sex-related genes DM-W and DMRT1 in African clawed frogs (Xenopus). Evolution 65 698 712
39. StockMHornAGrossenCLindtkeDSermierR 2011 Ever-young sex chromosomes in European tree frogs. PLoS Biol 9 e1001062 doi:10.1371/journal.pbio.1001062
40. LeeKHYamaguchiARashidHKadomuraKYasumotoS 2009 Estradiol-17ß treatment induces intersexual gonadal development in the pufferfish, Takifugu rubripes. Zool Sci 26 639 645
41. LaurieCCNickersonDAAndersonADWeirBSLivingstonRJ 2007 Linkage disequilibrium in wild mice. PLoS Genet 3 e144 doi:10.1371/journal.pgen.0030144
42. SlateJPembertonJ 2007 Admixture and patterns of linkage disequilibrium in a free-living vertebrate population. J Evo Biol 20 1415 1427
43. SaitohKChenWJ 2008 Reducing cloning artifacts for recovery of allelic sequences by T7 endonuclease I cleavage and single re-extension of PCR products - A benchmark. Gene 423 92 95
44. BarrettJCFryBMallerJDalyMJ 2005 Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21 263 265
45. LarkinMBlackshieldsGBrownNChennaRMcGettiganP 2007 Clustal W and Clustal X version 2.0. Bioinformatics 23 2947 2948
46. OhtaniMMiyadaiT 2011 Functional analysis of fish BCL-6 and Blimp-1 in vitro: Transcriptional repressors for B-cell terminal differentiation in fugu (Takifugu rubripes). Mol Immunol 48 818 825
47. SchwedeTKoppJGuexNPeitschMC 2003 SWISS-MODEL: an automated protein homology-modeling server. Nucleic Acids Res 31 3381 3385
48. GuexNPeitschMC 1997 SWISS-MODEL and the Swiss-Pdb Viewer: an environment for comparative protein modeling. Electrophoresis 18 2714 2723
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 7
Nejčtenější v tomto čísle
- Guidelines for Genome-Wide Association Studies
- The Role of Rice HEI10 in the Formation of Meiotic Crossovers
- Identification of Chromatin-Associated Regulators of MSL Complex Targeting in Dosage Compensation
- GWAS Identifies Novel Susceptibility Loci on 6p21.32 and 21q21.3 for Hepatocellular Carcinoma in Chronic Hepatitis B Virus Carriers