
Direct Visualization by Cryo-EM of the Mycobacterial Capsular Layer: A Labile Structure Containing ESX-1-Secreted Proteins
The cell envelope of mycobacteria, a group of Gram positive bacteria, is composed of a plasma membrane and a Gram-negative-like outer membrane containing mycolic acids. In addition, the surface of the mycobacteria is coated with an ill-characterized layer of extractable, non-covalently linked glycans, lipids and proteins, collectively known as the capsule, whose occurrence is a matter of debate. By using plunge freezing cryo-electron microscopy technique, we were able to show that pathogenic mycobacteria produce a thick capsule, only present when the cells were grown under unperturbed conditions and easily removed by mild detergents. This detergent-labile capsule layer contains arabinomannan, α-glucan and oligomannosyl-capped glycolipids. Further immunogenic and proteomic analyses revealed that Mycobacterium marinum capsule contains high amounts of proteins that are secreted via the ESX-1 pathway. Finally, cell infection experiments demonstrated the importance of the capsule for binding to cells and dampening of pro-inflammatory cytokine response. Together, these results show a direct visualization of the mycobacterial capsular layer as a labile structure that contains ESX-1-secreted proteins.
Published in the journal:
. PLoS Pathog 6(3): e32767. doi:10.1371/journal.ppat.1000794
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1000794
Summary
The cell envelope of mycobacteria, a group of Gram positive bacteria, is composed of a plasma membrane and a Gram-negative-like outer membrane containing mycolic acids. In addition, the surface of the mycobacteria is coated with an ill-characterized layer of extractable, non-covalently linked glycans, lipids and proteins, collectively known as the capsule, whose occurrence is a matter of debate. By using plunge freezing cryo-electron microscopy technique, we were able to show that pathogenic mycobacteria produce a thick capsule, only present when the cells were grown under unperturbed conditions and easily removed by mild detergents. This detergent-labile capsule layer contains arabinomannan, α-glucan and oligomannosyl-capped glycolipids. Further immunogenic and proteomic analyses revealed that Mycobacterium marinum capsule contains high amounts of proteins that are secreted via the ESX-1 pathway. Finally, cell infection experiments demonstrated the importance of the capsule for binding to cells and dampening of pro-inflammatory cytokine response. Together, these results show a direct visualization of the mycobacterial capsular layer as a labile structure that contains ESX-1-secreted proteins.
Introduction
Mycobacteria are the causative agent of tuberculosis and other chronic diseases such as leprosy and causes about 1.7 million deaths annually. The interaction of Mycobacterium tuberculosis with its macrophage host cell is largely dictated by its unique cell envelope components that are able to elicit immuno-modulatory responses similar to that of the pathogen [1].
The mycobacterial cell envelope has a complex structure composed of a typical phospholipid bilayer plasma membrane (PM), an outer membrane and an outermost layer known as the capsule in the case of pathogenic species [2]. The mycobacterial outer membrane henceforth referred to as ‘mycomembrane’ is mainly composed of long chain (C60-C90) mycolic fatty acids with free intercalating glycolipids and is covalently linked to the arabinogalactan-peptidoglycan layer [3],[4]. The chemical nature of mycomembrane has been examined in detail [4] and recent cryo-transmission electron microscopy (EM) data shows that it organizes into a structure analogous to the outer membrane of Gram-negative bacteria [5],[6]. The structural organization of this highly insoluble matrix is invariable among different mycobacterial species and is responsible for the low permeability of the mycobacterial cell envelope [4].
The capsule is visible as an electron transparent zone (ETZ) surrounding the mycobacterial cell envelope [2],[7] in conventional EM preparations [8],[9]. This layer was not, however, observed by cryo-EM [5],[6], questioning somehow its existence. This layer, mainly composed of polysaccharides, proteins and small amounts of lipids, is considered to have a different molecular composition in pathogenic and non-pathogenic species [2],[10],[11]. The presence of the capsular layer is influenced by the culturing conditions used; laboratory cultures commonly grown in the presence of detergent with agitation to prevent clumping [12] usually shed the capsule into the medium [13],[14].
In the present study, we adopt a close to native-state approach to study the mycobacterial cell envelope and demonstrate that mycobacteria express a distinctive outer layer. We have characterized the mycobacterial capsule layer by immunological and proteomic analyses and showed that some mycobacteria have ESX-1 secreted proteins in their capsule layer. ESX-1 encodes a type VII secretion system that mediates the secretion of potent T cell antigens such as ESAT-6 (EsxA) and CFP-10. Furthermore, the presence of a capsule not only enhanced the Mycobacterium-macrophage interaction but also dampened the pro-inflammatory cytokine response.
Results/Discussion
Cell envelope organization of mycobacterial cells
The ultra-structure of the mycobacterial cell envelope was investigated using 30 nm vitreous sections, as previously described [15]. As a control for structural preservation of the envelope morphology, we also examine vitreous sections of the Gram-negative bacterium Shigella flexneri and the Gram-positive bacterium Staphylococcus epidermidis. The width of the PM and outer membrane or cell wall in both species are consistent with previous results [16],[17] (Figure S1, Table S1). The vitreous sections of four mycobacterial species [M. tuberculosis, M. marinum, Mycobacterium bovis Bacille Calmette-Guérin (BCG) and Mycobacterium smegmatis] examined appeared similar and show a well preserved cell envelope composed of a PM and a mycomembrane with a bilayer profile (Figure S1C, S2 and S3). Although the intensity of the mycobacterial PM is similar to that of S. flexneri and S. epidermidis, the width (≈7 nm) is at least 16% wider (Table S1). The mycomembrane, with an apparent thickness of ≈8.3 nm is also thicker than the 6.8 nm thick outer membrane of S. flexneri. This slight increase in thickness is similar to recently published data [5],[6].
Cryo-electron tomography analysis was performed to determine the 3D architecture of the envelope (Figure S1D-F). In the reconstructed slices of M. smegmatis cell envelope, two additional layers are present in the periplasmic space. These layers are similar to those observed as L1 and L2 in [5] and as the granular layer and the medial wall zone referred to in [6]. The L1 layer appears immediately after the plasma membrane while the L2 layer appears closely apposed to the mycomembrane and may possibly be the peptidoglycan/arabinogalactan matrix. Taken together, these findings show that the cell envelope morphology of the mycobacteria is structurally related to, but more complex than that of the Gram-negative bacteria.
Visualization of the mycobacterial capsular layer and its disruption by Tween and agitation
The capsule, being a substantial component of the cell envelope, should be visible in vitreous sections as an extra layer extending from the mycomembrane. Yet, in our analyses and previous studies [5],[6], such a layer is not visible (Figure S1C, S3). We reasoned that the detection of this layer may have been obscured by the presence of the dextran cryoprotectant, which may have an electron density similar to that of the capsule. Alternatively, the capsule might have been removed by growing the cells in the presence of detergent and agitation in order to prevent clumping [2],[13],[14].
To investigate this, mycobacterial cells were cultured with or without Tween-80 and agitation and subsequently frozen in a close to native-state by the plunge freezing method [18] for direct visualization by EM (Figure 1). This method does not rely on the use of cryoprotectants and allows intact cells to be frozen in their medium of culture. The Gram-negative bacterium S. flexneri was used as a control (Figure 1A). When grown without any perturbation, all mycobacterial species examined, showed a thick outermost capsule-like layer (Figure 1C to 1F). To our knowledge, this is the first time this layer is visualized in a close to native state surrounding both pathogenic and nonpathogenic mycobacterial species. In comparison, cells grown with perturbation show this layer to be partially or completely removed (Figure 1B), meaning that growing mycobacteria under routine culturing conditions involving perturbation promotes shedding of this layer.

To demonstrate that this outermost layer is distinct from the mycomembrane, we sought to localize OmpATb porin known to be present in the mycomembrane [19] by labeling fixed cryo-sections of M. smegmatis cells over-expressing this protein [20] with anti-OmpATb serum and probed with protein-A attached to 10 nm gold. When grown in the presence of perturbation, the mycomembrane was efficiently labeled with the antibody (Figure 2A). In comparison, cells grown in the absence of the detergent maintain the outermost layer, though the OmpATb antibody only specifically labeled the underlying mycomembrane (Figure 2B). Parental M. smegmatis strain, naturally devoid of OmpATb, cultured under unperturbed state lacks any labeling, demonstrating the specificity of this antibody (Figure 2C). These results indicate that the outermost layer removed by perturbation is distinct and not part of the mycomembrane.

Characterization of the mycobacterial capsular layer by immuno gold-EM
To confirm that the observed outermost layer is indeed the capsule, immuno gold-EM analysis of whole bacteria using antisera against known capsular components was performed. Whole bacteria were immobilized on EM grids and incubated with specific antibodies and gold probes. This method also allowed us to visualize and quantify the different components. Since α-glucan is described to be the major capsular polysaccharide [2],[11], we first investigated this using an α-glucan-specific monoclonal antibody [21]. The bacterial surface of cells grown in unperturbed state was distinctly labeled homogenously with this antibody (Figure 3), whereas cells grown in perturbed state showed weak or no labeling. Fixation of the cells was important for efficient labeling, since the same unperturbed culture without any fixation showed reduced or weak labeling. Non-acylated arabinomannan (AM) is also a described component of the capsule [11],[22]. Using antibodies directed against arabinomannan structural motifs, our experiments confirmed the presence of AM in the capsule (Figure S4) Subsequently, we tried to localize the glycolipid phosphatidylinositol mannosides (PIMs), which is possibly associated with the capsule [23]. For this we used an antibody that on M. smegmatis specifically recognizes the PIM6 epitope, as it does not interact with M. smegmatis pimE mutant (Figure 3F) [24],[25]. The antibody labeled the surface of intact cells very efficiently, supporting the presence of specific PIM epitopes on M. smegmatis and possibly other mannoconjugates and PIM on the pathogenic mycobacteria except for M. marinum which is poorly labeled with this antibody. Importantly, all of the antibodies significantly labeled less when the bacteria were grown agitated in the presence of detergent (Figure 3A, 3D and S4). Agitation in the absence of detergent contributes to the erosion of the capsule albeit lesser than when detergent is also included, conversely, the addition of detergent alone also shows less capsular disruption, thus the effect is additive.

In addition to glycans and glycolipids, the capsule also contains proteins [2],[10], although these are generally not well described. One putative capsular protein recently described by Carlsson et al [26] is the ESX-1-associated protein EspE [formerly known as Mh3864 [27]], which together with EspB is also secreted by ESX-1 encoded type VII secretion system [26],[28]. To investigate EspE localization, whole cells were labeled with anti-EspE antibody which homogenously labeled the surface of M. marinum and M. tuberculosis. A preference of EspE for the polar region as previously reported [26] was not observed (Figure 3G-I). The localization of EspE in a M. marinum strain (Fas 3.1) with a transposon insertion in eccCb1 [formerly known as Mh1784 [27]] which blocks ESX-1-dependent secretion (van der Sar et al, in preparation), showed no specific labeling, demonstrating both the specificity of the antibody used and the ESX-1 dependence of EspE export. Finally, EspE labeling was not strongly affected by Tween-80, suggesting that this protein is more tightly associated with the cell envelope.
Capsule of M. marinum contains secretion specific proteins
To screen for specific (ESX-secreted) proteins within the capsule for which no antibodies are available, we analyzed capsular extracts by liquid chromatography mass spectrometry (LC-MS). For this, capsular material was isolated from cells grown under unperturbed conditions by treating the cells with the mild detergents Tween-80 (1%) or Genapol-X080 (0.25%) (Figure 4). First, we analyzed the isolated fractions and the cell pellet for the presence of capsular α-glucan by a spot-blot assay. Consistent with our EM data, both M. marinum and M. smegmatis cells contained more α-glucan when they were grown in the absence of Tween-80 (Figure 4A and S5, respectively). Furthermore, spot-blot analysis showed that α-glucan was extracted by these mild detergents. These fractions were subsequently analyzed by SDS-PAGE. Extracts from bacteria grown in the absence of detergent repeatedly contained higher amounts of proteins, which indicate that their presence depends on an intact capsule (Figure 4B). In addition, more proteins were extracted by Genapol as compared to Tween.

The Genapol-extracted fractions from bacteria grown in the absence of Tween-80 were analyzed by LC-MS (Table 1). The highest number of peptides found in the M. marinum extract belonged to EspE, whose predicted molecular weight of 40 kDa corresponds to the size of the most abundant protein in the extract (Figure 4B). Interestingly, other ESX-1-associated proteins such as EspB, EspF (Mh3865), EspK (Mh3879c), ESAT-6 (EsxA), CFP-10 and PPE68_1 were also present in high amounts. In addition, several other PE and PPE proteins were found in the Genapol extracts. OmpA was not detected in the Genapol-extracted material, confirming that this treatment did not extract proteins from the mycomembrane. These data show that M. marinum has a specific set of proteins within its capsule.

The situation was different for M. smegmatis and M. tuberculosis. The capsular extracts of these two species did not show any major protein bands (Figure 4B). This observation was confirmed by LC-MS analysis, which showed that the major proteins were cytosolic proteins such as GroEL2, DnaK and translation elongation factor Tu (Table S2 and S3). The presence of some of these proteins in the capsule was also recently described by Hickey et al. [29]. In addition to these predominantly cytosolic proteins only very small amounts of putative capsular proteins, such as the ESX-1-associated proteins, were identified by LC-MS (data not shown). This could indicate that the capsular proteins of M. smegmatis and M. tuberculosis might be more tightly associated with the cell envelope than in M. marinum and are therefore missed during analysis.
The major capsular proteins of M. marinum are secreted via ESX-1
The capsule of M. marinum contains a high amount of ESX-1 associated protein. To investigate whether these proteins depend on ESX-1 for their localization, Genapol extracted proteins of the M. marinum Fas 3.1 eccCb1 mutant were analyzed (Figure 4C). The protein pattern of the ESX-1 mutant differed considerably from the wild-type protein pattern. Importantly, the 40 kDa product corresponding to EspE was not detectable in the capsular extract of the ESX-1 mutant (Figure 4C and D), suggesting that the capsular localization of EspE depends on an active ESX-1 secretion system in M. marinum as previously reported [26]. This result was also confirmed by LC-MS analysis (Table 1) and shows that EspF, EspK and the PPE proteins associated with the ESX-1 pathway are dependent on an active ESX-1 system for capsular localization and are therefore new putative ESX-1 substrates. The presence of other PE and PPE proteins in the capsule was not affected by the ESX-1 mutation, which is in agreement with the recent observation that many of these proteins, including the capsule PPE protein MMAR_1402, depend on another secretion system, ESX-5 for their export [30].
Capsular layer promotes binding to macrophages and inhibits pro-inflammatory cytokine response
As demonstrated above, growing mycobacteria in the presence of detergent with agitation, promotes capsular shedding and thus may influence the biological characteristics of the bacteria. To investigate this issue and to potentially gain insight into the biological relevance of the capsule, M. bovis BCG was grown under unperturbed conditions after which the bacteria were treated with 1% Tween-80 to remove the capsule. First, we investigated the consequence of capsule removal on the ability of the bacteria to bind to human monocyte-derived macrophages. As shown in Figure 5A, removal of the capsule significantly reduced bacterial binding at all tested multiplicities of infection (MOIs), suggesting that the presence of the capsule promoted association to the macrophages. Next, we investigated whether the presence of a capsule may also differentially modulate the macrophage pro-inflammatory cytokine responses. As shown in Figure 5B, non-detergent-treated M. bovis BCG induced significantly lower amounts of IL-12p40, IL-6, and TNFα (∼30%) as compared to the detergent-treated bacteria at both MOIs tested. The only exception was IL-6, for which the difference at MOI 8 was non-significant. Reduced cytokine induction by untreated (encapsulated) M. bovis BCG was observed in both resting macrophages (Figure 5B) and in macrophages primed with LPS (data not shown). Interestingly, treatment with detergent did not differentially affect the macrophage response for non-pathogenic M. smegmatis (Figure 5C). Overall, these findings demonstrate that at least for M. bovis BCG the capsule is involved in facilitating macrophage infection and, simultaneously, plays a role in dampening the pro-inflammatory macrophage response. Furthermore, these data imply that growing mycobacteria in the presence of detergent, as is done under many standard laboratory circumstances, may have important consequences for the biological characteristics of these bacteria.

Summary and biological implications
In this study we used a plunge freezing method [18] that avoids artifacts introduced by chemical fixation and thus gives a close to native estimation of the thickness for these in vitro grown bacteria. We demonstrated the presence of the capsular layer in the cell envelope of both pathogenic and nonpathogenic mycobacterial species and determined its thickness to be around 30 nm (Table S1), taking into account the instability of the capsule in vitro; both groups have been previously shown to differ in the amount of extracellular material found in static and non-detergent-treated medium, which is interpreted as representing part of the capsule shed from growing cells [10],[11]. One possibility of circumventing this instability is to measure this layer inside phagocytic cells in the vitreous state. It is however currently very difficult to freeze large mammalian cells under physiological conditions in a vitreous state [31].
The detection of capsular components α-glucan, arabinomannan, and oligo - and poly-mannosylated compounds on the surface of whole cells by immuno gold-EM is in agreement with chemical data about the nature of the capsule [4],[10],[11]. The presence of PIMs exposed at the surface of mycobacteria was already demonstrated [32],[33],[34], however here we showed that PIM, α-glucan, and ESX secreted proteins are accessible to antibodies on intact bacteria. Mycobacterial α-glucan is implicated in the regulation of phagocytosis [35],[36], modulation of immune response [1],[21], and persistence of infection [37]. In this study we have shown that binding to macrophages and a dampening of the pro-inflammatory cytokine response is observed for encapsulated M. bovis BCG, which suggests that components of the capsule, such as glycans, play a role in immune response.
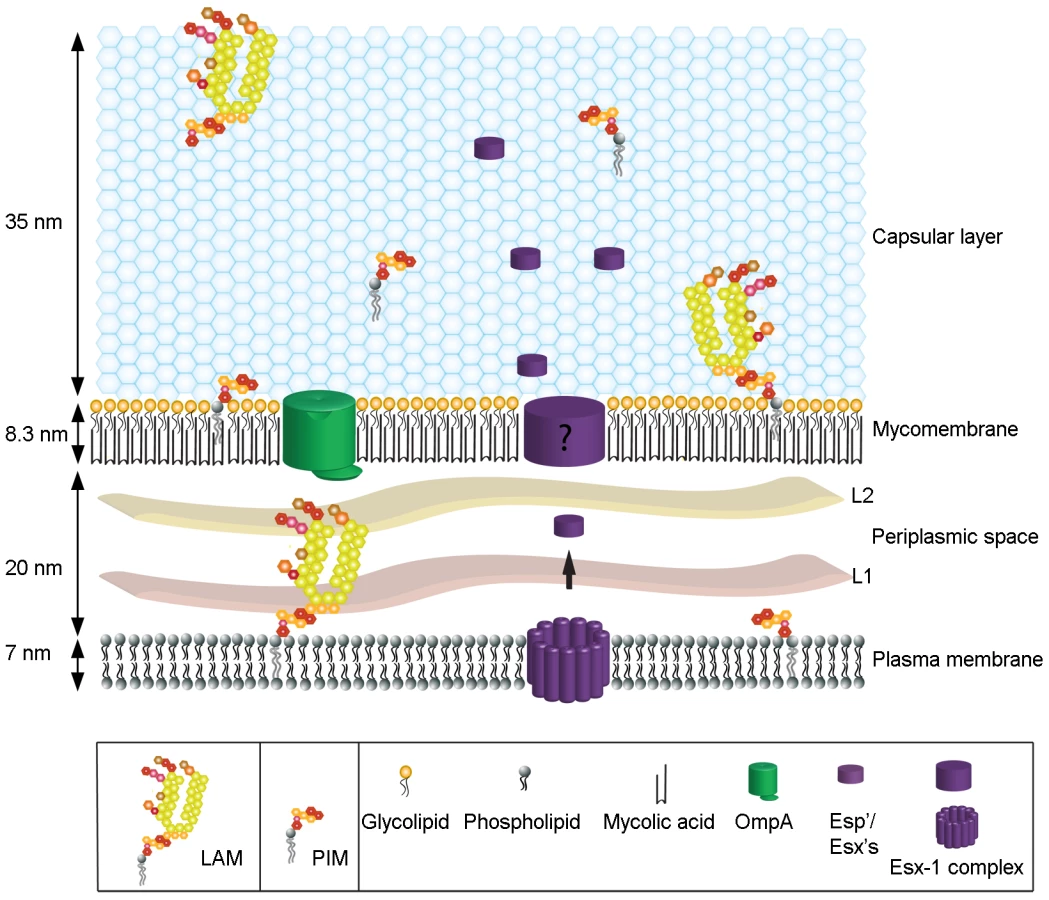
The identification of many ESX-5 and ESX-1 secreted proteins (ESAT-6 and CFP-10, EspB, EspE, EspF, EspK and an ESX-1 associated PPE) in the capsule, a number of which have not been identified previously as being secreted via ESX-1 (EspF, EspK and PPE68_1) was surprising. These proteins were only detectable in the detergent-labile capsule fraction of M. marinum. Some ESX-1-secreted proteins have an essential role in the interaction with the host [38],[39],[40], and more specifically in bacterial translocation from the phagosome into the cytosol [41]. The fact that EspE can only be partly removed by Tween-80, suggests a distinction between proteins that are freely associated with the capsule and those more tightly associated with the rest of the cell envelope. In this context, these ESX-1 associated proteins may form a surface-exposed macromolecular structure through (and/or within) the capsule, specialized for the interaction with the host (Figure 6). Together, we showed that the mycobacterial capsule contains various components that manipulate the host, making this layer an attractive target for vaccine and drug development.
Materials and Methods
Bacterial strains and growth conditions
M. tuberculosis 6020 strain [42], M. bovis BCG strain Copenhagen, M. smegmatis mc2155, M. smegmatis-OmpATb [20], M. marinum E11 strain and the E11 fas 3.1 eccCb1 mutant (van der Sar et al, in preparation) were grown at 37°C (or 30°C for M. marinum) in Middlebrook 7H9 media (Difco) supplemented with 10% oleic acid-albumin-dextrose catalase (OADC) (BBL) with or without 0.05% Tween 80 where applicable. For the growth of M. smegmatis-OmpATb and M. bovis BCG expressing dsRed, 25 µg and 50 µg mL−1 kanamycin and 50 µg mL−1 hygromycin were added, respectively. S. flexneri ipaC strain SF621 [43] was grown in tryptic casein soy broth (Sigma) at 37°C. S. epidermidis was grown in Luria-Bertani broth (Difco) at 37°C.
Cryo-EM
Vitrification, vitreous sectioning and tomography is as described in Supplementary information (SI) Text and [44]. For plunge freezing, bacteria were taken directly from culture medium for processing without further centrifugation. A 4 µl droplet of various mycobacteria sample were applied to a glow-discharge quantifoil copper grids (Quantifoil Micro Tools, Jena, Germany) mounted in an environmentally controlled chamber at 100% humidity, blotted and frozen in vitreous ice by plunging into liquid ethane using the vitrobot (FEI). Grids were transferred to a Gatan model 626 cryoholder (Gatan, Pleasanton, CA) under liquid nitrogen and inserted into a Tecnai 12 (FEI, Eindhoven, Netherlands) operating at 120 kV. The vitreous state of the preparation was confirmed by electron diffraction. Low-dose images, with exposures between 10 and 20 electrons per Å2 and under-focus values of 2 to 4 µm were recorded with a 4096×4096 pixel CCD camera (Gatan) at ×18,000 −×23,000 magnification.
Immuno gold-EM
Bacteria cells were fixed by addition of an equal volume of 0.4M PHEM buffer containing 4% paraformaldehyde and 0.4% glutaraldehyde to the culture and incubating for 2 hours at room temperature [41].
For whole mount cells, samples were taken directly from the fixed culture without pelleting and incubated for 5 minutes on carbon coated formvar grids. The rabbit antibody used is OmpATb [19]. The following murine IgM monoclonal antibodies (Mabs) were used: anti-Ara6 (F30-5) recognizes Ara6-motive in the arabinan domain of (L)AMs [45],[46]; anti-ManLAM (55.92.1A1) binds the mannosyl caps on mannosyl-capped (L)AMs [46]; anti-PIM6 (F183-24) recognizes terminal α(1,2)-linked mannosyl residues as present in PIM6 and the mannosyl caps on ManLAM [25],[45]. In M. smegmatis [which lacks the mannosyl-caps on (L)AM] F183-24 specifically recognizes PIM6 with no reactivity to other components [25]. The Mab against capsular α-glucans was generated by O. Baba [47]. Immuno-labeling is essentially as described [see Text S1 and [41]]. Labeled whole mount cells were observed without further staining with a Tecnai 12 or CM 10 electron microscope (FEI, Eindhoven, Netherlands).
Cryo-sectioning and immuno gold labeling of fixed cells is essentially as described in Text S1 and [41].
Capsule extraction
Mycobacteria were grown under conditions mentioned above to an OD600 of ∼1, diluted 100 times in 7H9 without supplement or Tween-80, but with 0.2% dextrose and grown to exponential phase. Bacteria were harvested by centrifugation and washed 3 times in PBS. Bacteria were resuspended in PBS and incubated with 1% Tween-80 or 0.25% Genapol X-080 for 30 min. at room temperature before centrifugation to separate extracted fractions from bacteria cells. To determine the presence of α-glucan the extracted fractions were spotted on a nitrocellulose membrane, the membrane was dried for 1 hour at 80°C, blocked by 5% skimmed milk in PBS-T overnight and immuno-labeled using an anti-α-glucan monoclonal antibody. For analysis of the protein content, the extracted fraction was concentrated by TCA precipitation and analyzed via SDS-PAGE and Coomassie staining or Western blotting using anti-EspE [26].
Sample preparation for mass spectrometric analysis
Protein lanes from Coomassie stained SDS-PAGE gels were excised and prepared for LC-MS analysis as described in Text S1.
Preparation of mycobacteria for macrophage-binding and stimulation assays
M. bovis BCG, M. bovis BCG expressing dsRed, or M. smegmatis were grown in Middlebrook 7H9 broth (Difco) supplemented with 10% ADC enrichment in the absence of Tween-80. Exponentially growing bacteria were collected and washed once with 50 ml PBS. The bacteria were resuspended in 20 ml PBS, split into two 10 ml fractions, and incubated with/without 1% Tween-80 for 30 min with rotation at room temperature. The treated and untreated bacteria were washed twice with 25 mL Tris-buffered saline supplemented with 1 mM MgCl2 and 2 mM CaCl2 (TSM) containing 0.5% human serum albumin (HSA), passed through a 5 µm filter (Millipore) for declumping (bacterial suspensions were first centrifuged for 10 min at 200×g to prevent filter-clogging), and diluted with TSM/0.5% HSA to the appropriate optical density. Colony forming units were determined by plating the suspensions on 7H10 agar supplemented with 10% Middlebrook OADC.
Macrophage binding assay
M. bovis BCG expressing dsRed was added to 5×105 macrophages [generated as described in Text S1 and ref.[21]] in 100 µL TSM/0.5% HSA at a MOI of 8, 2, or 0.5. After 45 min at 4°C, cells were washed twice with TSM/0.5% HSA and the percentage of fluorescent cells was determined using a FACScan Analytic Flow Cytometer (Becton Dickinson). Data was analyzed using manufacturer's software (CellQuest version 3.1f).
Macrophage stimulation assay
Macrophages were released by trypsiniation and resuspended in culture medium (containing 50 U mL−1 GM-CSF) at a concentration of 1.25×106 cells mL−1. Eighty µl cell suspension (1×105 cells) was transferred to a sterile 96-well U-bottom plate (Greiner) and left for 16 h (37°C, 5% CO2) to allow cells to adhere. Macrophages were stimulated with M. bovis BCG or M. smegmatis (treated with or without 1% Tween-80) in the absence or presence of 20 ng mL−1 lipopolysaccharide (LPS) (from Salmonella enterica serotype abortus equi (Sigma-Aldrich L5886). Unstimulated cells served as controls in all experiments. Culture supernatants were harvested after 24 h of incubation (37°C, 5% CO2) by centrifugation and stored at −80°C for cytokine measurements using an enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's instructions (Invitrogen). Data were statistically analyzed using a Student's t test (two-tailed, two-sample equal variance). Differences were considered to be significant when p<0.05.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. GagliardiMC
LemassuA
TeloniR
MariottiS
SargentiniV
2007 Cell wall-associated alpha-glucan is instrumental for Mycobacterium tuberculosis to block CD1 molecule expression and disable the function of dendritic cell derived from infected monocyte. Cell Microbiol 9 2081 2092
2. DaffeM
EtienneG
1999 The capsule of Mycobacterium tuberculosis and its implications for pathogenicity. Tubercle Lung Dis 79 153 169
3. MinnikinDE
1982 Lipids: complex lipids, their chemistry, biosynthesis and roles.The Biology of Mycobacteria.
RatledgeC
StandfordJ
The Biology of Mycobacteria London Academic 95 184
4. BrennanPJ
CrickDC
2007 The cell-wall core of Mycobacterium tuberculosis in the context of drug discovery. Curr Top Med Chem 7 475 488
5. HoffmannC
LeisA
NiederweisM
PlitzkoJM
EngelhardtH
2008 Disclosure of the mycobacterial outer membrane: cryo-electron tomography and vitreous sections reveal the lipid bilayer structure. Proc Natl Acad Sci U S A 105 3963 3967
6. ZuberB
ChamiM
HoussinC
DubochetJ
GriffithsG
2008 Direct visualization of the outer membrane of mycobacteria and corynebacteria in their native state. J Bacteriol 190 5672 5680
7. ChapmanGB
HanksJH
WallaceJH
1959 An electron microscope study of the disposition and fine structure of Mycobacterium lepraemurium in mouse spleen. J Bacteriol 77 205 211
8. FrehelC
RyterA
RastogiN
DavidH
1986 The electron-transparent zone in phagocytized Mycobacterium avium and other mycobacteria: formation, persistence and role in bacterial survival. Ann Inst Pasteur Microbiol 137B 239 257
9. PaulTR
BeveridgeTJ
1994 Preservation of surface lipids and determination of ultrastructure of Mycobacterium kansasii by freeze-substitution. Infect Immun 62 1542 1550
10. Ortalo-MagneA
DupontMA
LemassuA
AndersenAB
GounonP
1995 Molecular composition of the outermost capsular material of the tubercle bacillus. Microbiology 1609 1620 141 (UK)
11. LemassuA
Ortalo-MagneA
BardouF
SilveG
LaneelleMA
1996 Extracellular and surface-exposed polysaccharides of non-tuberculous mycobacteria. Microbiology 1513 1520 142 (UK)
12. WrightEL
PourshafieM
BarrowWW
1992 Mycobacterium avium rough-to-smooth colony conversion resulting from growth in Tween 80 without presence of type-specific glycopeptidolipid antigens. FEMS Microbiol Lett 77 209 216
13. SchwebachJR
CasadevallA
SchneersonR
DaiZ
WangX
2001 Expression of a Mycobacterium tuberculosis arabinomannan antigen in vitro and in vivo. Infect Immun 69 5671 5678
14. SchwebachJR
Glatman-FreedmanA
Gunther-CumminsL
DaiZ
RobbinsJB
2002 Glucan is a component of the Mycobacterium tuberculosis surface that is expressed in vitro and in vivo. Infect Immun 70 2566 2575
15. ZhangP
BosE
HeymannJ
GnaegiH
KesselM
2004 Direct visualization of receptor arrays in frozen-hydrated sections and plunge-frozen specimens of E. coli engineered to overproduce the chemotaxis receptor Tsr. J Microsc 216 76 83
16. MatiasVR
Al AmoudiA
DubochetJ
BeveridgeTJ
2003 Cryo-transmission electron microscopy of frozen-hydrated sections of Escherichia coli and Pseudomonas aeruginosa. J Bacteriol 185 6112 6118
17. DubochetJ
McDowallAW
MengeB
SchmidEN
LickfeldKG
1983 Electron microscopy of frozen-hydrated bacteria. J Bacteriol 155 381 390
18. DubochetJ
AdrianM
ChangJJ
HomoJC
LepaultJ
1988 Cryo-electron microscopy of vitrified specimens. Q Rev Biophys 21 129 228
19. SenaratneRH
MobasheriH
PapavinasasundaramKG
JennerP
LeaEJ
1998 Expression of a gene for a porin-like protein of the OmpA family from Mycobacterium tuberculosis H37Rv. J Bacteriol 180 3541 3547
20. AlahariA
SaintN
CampagnaS
MolleV
MolleG
2007 The N-terminal domain of OmpATb is required for membrane translocation and pore-forming activity in mycobacteria. J Bacteriol 189 6351 6358
21. GeurtsenJ
ChedammiS
MestersJ
CotM
DriessenNN
2009 Identification of mycobacterial alpha-glucan as a novel ligand for DCSIGN:involvement of mycobacterial capsular polysaccharides in host immune modulation. J Immunol 183 5221 5231
22. Ortalo-MagneA
AndersenAB
DaffeM
1996 The outermost capsular arabinomannans and other mannoconjugates of virulent and avirulent tubercle bacilli. Microbiology 927 935 142 (UK)
23. Ortalo-MagneA
LemassuA
LaneelleMA
BardouF
SilveG
1996 Identification of the surface-exposed lipids on the cell envelopes of Mycobacterium tuberculosis and other mycobacterial species. J Bacteriol 178 456 461
24. MoritaYS
SenaCB
WallerRF
KurokawaK
SerneeMF
2006 PimE is a polyprenol-phosphate-mannose-dependent mannosyltransferase that transfers the fifth mannose of phosphatidylinositol mannoside in mycobacteria. J Biol Chem 281 25143 25155
25. DriessenNN
UmmelsR
MaaskantJJ
GurchaSS
BesraGS
2009 Role of phosphatidylinositol mannosides in the interaction between mycobacteria and DC-SIGN. Infect Immun 77 4538 4547
26. CarlssonF
JoshiSA
RangellL
BrownEJ
2009 Polar localization of virulence-related Esx-1 secretion in mycobacteria. PLoS Path 5 e1000285 doi:10.1371/journal.ppat.1000285
27. BitterW
HoubenEN
BottaiD
BrodinP
BrownEJ
2009 Systematic genetic nomenclature for type VII secretion systems PLoS Path 5 e1000507 doi:10.1371/journal.ppat.1000507
28. McLaughlinB
ChonJS
MacGurnJA
CarlssonF
ChengTL
2007 A mycobacterium ESX-1-secreted virulence factor with unique requirements for export. PLoS Path 3 e105 doi:10.1371/journal.ppat.0030105
29. HickeyTB
ThorsonLM
SpeertDP
DaffeM
StokesRW
2009 Mycobacterium tuberculosis Cpn60.2 and DnaK are located on the bacterial surface, where Cpn60.2 facilitates efficient bacterial association with macrophages. Infect Immun 77 3389 3401
30. AbdallahAM
VerboomT
WeerdenburgEM
Gey van PittiusNC
MahashaPW
2009 PPE and PE_PGRS proteins of Mycobacterium marinum are transported via the type VII secretion system ESX-5. Mol Microbiol 73 329 340
31. PiersonJ
SaniM
TomovaC
GodsaveS
PetersPJ
2009 Toward visualization of nanomachines in their native cellular environment. Histochem Cell Biol 132 253 262
32. HunterSW
BrennanPJ
1990 Evidence for the presence of a phosphatidylinositol anchor on the lipoarabinomannan and lipomannan of Mycobacterium tuberculosis. J Biol Chem 265 9272 9279
33. OzanneV
Ortalo-MagneA
VercelloneA
FournieJJ
DaffeM
1996 Cytometric detection of mycobacterial surface antigens: exposure of mannosyl epitopes and of the arabinan segment of arabinomannans. J Bacteriol 178 7254 7259
34. PitarqueS
Larrouy-MaumusG
PayreB
JacksonM
PuzoG
2008 The immunomodulatory lipoglycans, lipoarabinomannan and lipomannan, are exposed at the mycobacterial cell surface. Tuberculosis 88 560 565
35. CywesC
HoppeHC
DaffeM
EhlersMR
1997 Nonopsonic binding of Mycobacterium tuberculosis to complement receptor type 3 is mediated by capsular polysaccharides and is strain dependent. Infect Immun 65 4258 4266
36. StokesRW
Norris-JonesR
BrooksDE
BeveridgeTJ
DoxseeD
2004 The glycan-rich outer layer of the cell wall of Mycobacterium tuberculosis acts as an antiphagocytic capsule limiting the association of the bacterium with macrophages. Infect Immun 72 5676 5686
37. SambouT
DinadayalaP
StadthagenG
BariloneN
BordatY
2008 Capsular glucan and intracellular glycogen of Mycobacterium tuberculosis: biosynthesis and impact on the persistence in mice. Mol Microbiol 70 762 774
38. PymAS
BrodinP
BroschR
HuerreM
ColeST
2002 Loss of RD1 contributed to the attenuation of the live tuberculosis vaccines Mycobacterium bovis BCG and Mycobacterium microti. Mol Microbiol 46 709 717
39. GaoLY
GuoS
McLaughlinB
MorisakiH
EngelJN
2004 A mycobacterial virulence gene cluster extending RD1 is required for cytolysis, bacterial spreading and ESAT-6 secretion. Mol Microbiol 53 1677 1693
40. StanleySA
RaghavanS
HwangWW
CoxJS
2003 Acute infection and macrophage subversion by Mycobacterium tuberculosis require a specialized secretion system. Proc Natl Acad Sci U S A 100 13001 13006
41. van der WelN
HavaD
HoubenD
FluitsmaD
van ZonM
2007 M. tuberculosis and M. leprae translocate from the phagolysosome to the cytosol in myeloid cells. Cell 129 1287 1298
42. SambandamurthyVK
DerrickSC
JalapathyKV
ChenB
RussellRG
2005 Long-term protection against tuberculosis following vaccination with a severely attenuated double lysine and pantothenate auxotroph of Mycobacterium tuberculosis. Infect Immun 73 1196 1203
43. MenardR
SansonettiPJ
ParsotC
1993 Nonpolar mutagenesis of the ipa genes defines IpaB, IpaC, and IpaD as effectors of Shigella flexneri entry into epithelial cells. J Bacteriol 175 5899 5906
44. PiersonJ
FernandezJJ
BosE
AminiS
GnaegiH
2009 Improving the technique of vitreous cryo-sectioning for cryo-electron tomography: electrostatic charging for section attachment and an anti-contamination Box. J Struct Biol: In press
45. KolkAH
HoML
KlatserPR
EggelteTA
KuijperS
1984 Production and characterization of monoclonal antibodies to Mycobacterium tuberculosis, M. bovis (BCG) and M. leprae. Clin Exp Immunol 58 511 521
46. AppelmelkBJ
den DunnenJ
DriessenNN
UmmelsR
PakM
2008 The mannose cap of mycobacterial lipoarabinomannan does not dominate the Mycobacterium-host interaction. Cell Microbiol 10 930 944
47. BabaO
1993 Production of monoclonal antibody that recognizes glycogen and its application for immunohistochemistry. Kokubyo Gakkai Zasshi The Journal of the Stomatological Society, Japan 60 264 287
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2010 Číslo 3
- Farmakovigilanční studie perorálních antivirotik indikovaných v léčbě COVID-19
- Jak souvisí postcovidový syndrom s poškozením mozku?
- Měli bychom postcovidový syndrom léčit antidepresivy?
- 10 bodů k očkování proti COVID-19: stanovisko České společnosti alergologie a klinické imunologie ČLS JEP
Nejčtenější v tomto čísle
- Kaposi's Sarcoma-Associated Herpesvirus ORF57 Protein Binds and Protects a Nuclear Noncoding RNA from Cellular RNA Decay Pathways
- Endocytosis of the Anthrax Toxin Is Mediated by Clathrin, Actin and Unconventional Adaptors
- Perforin and IL-2 Upregulation Define Qualitative Differences among Highly Functional Virus-Specific Human CD8 T Cells
- Exoerythrocytic Parasites Secrete a Cysteine Protease Inhibitor Involved in Sporozoite Invasion and Capable of Blocking Cell Death of Host Hepatocytes