
Eight RGS and RGS-like Proteins Orchestrate Growth, Differentiation, and Pathogenicity of
A previous study identified MoRgs1 as an RGS protein that negative regulates G-protein signaling to control developmental processes such as conidiation and appressorium formation in Magnaporthe oryzae. Here, we characterized additional seven RGS and RGS-like proteins (MoRgs2 through MoRgs8). We found that MoRgs1 and MoRgs4 positively regulate surface hydrophobicity, conidiation, and mating. Indifference to MoRgs1, MoRgs4 has a role in regulating laccase and peroxidase activities. MoRgs1, MoRgs2, MoRgs3, MoRgs4, MoRgs6, and MoRgs7 are important for germ tube growth and appressorium formation. Interestingly, MoRgs7 and MoRgs8 exhibit a unique domain structure in which the RGS domain is linked to a seven-transmembrane motif, a hallmark of G-protein coupled receptors (GPCRs). We have also shown that MoRgs1 regulates mating through negative regulation of Gα MoMagB and is involved in the maintenance of cell wall integrity. While all proteins appear to be involved in the control of intracellular cAMP levels, only MoRgs1, MoRgs3, MoRgs4, and MoRgs7 are required for full virulence. Taking together, in addition to MoRgs1 functions as a prominent RGS protein in M. oryzae, MoRgs4 and other RGS and RGS-like proteins are also involved in a complex process governing asexual/sexual development, appressorium formation, and pathogenicity.
Published in the journal:
. PLoS Pathog 7(12): e32767. doi:10.1371/journal.ppat.1002450
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1002450
Summary
A previous study identified MoRgs1 as an RGS protein that negative regulates G-protein signaling to control developmental processes such as conidiation and appressorium formation in Magnaporthe oryzae. Here, we characterized additional seven RGS and RGS-like proteins (MoRgs2 through MoRgs8). We found that MoRgs1 and MoRgs4 positively regulate surface hydrophobicity, conidiation, and mating. Indifference to MoRgs1, MoRgs4 has a role in regulating laccase and peroxidase activities. MoRgs1, MoRgs2, MoRgs3, MoRgs4, MoRgs6, and MoRgs7 are important for germ tube growth and appressorium formation. Interestingly, MoRgs7 and MoRgs8 exhibit a unique domain structure in which the RGS domain is linked to a seven-transmembrane motif, a hallmark of G-protein coupled receptors (GPCRs). We have also shown that MoRgs1 regulates mating through negative regulation of Gα MoMagB and is involved in the maintenance of cell wall integrity. While all proteins appear to be involved in the control of intracellular cAMP levels, only MoRgs1, MoRgs3, MoRgs4, and MoRgs7 are required for full virulence. Taking together, in addition to MoRgs1 functions as a prominent RGS protein in M. oryzae, MoRgs4 and other RGS and RGS-like proteins are also involved in a complex process governing asexual/sexual development, appressorium formation, and pathogenicity.
Introduction
Signal transduction cascades are the primary means by which external cues are communicated to the nuclei of eukaryotic organisms including fungi. Heterotrimeric guanine-nucleotide binding protein (G-protein) signaling is one of the most important mechanisms by which eukaryotic cells sense extracellular signals and integrate them into intrinsic signal transduction pathways, such as the cyclic AMP (cAMP)-dependent signaling pathway. Heterotrimeric G-proteins are activated by the seven-transmembrane-spanning family of receptors [1]. Binding of signal ligands to such receptors promotes an exchange of GDP to GTP on the Gα subunit, which then triggers a reciprocal conformational change and dissociation from the Gβγ heterodimer [2]. Either Gα or Gβγ, or both, are then free to activate downstream target effectors such as phosphodiesterase, protein kinases, adenylyl cyclases, phospholipases, and ion channels [3]–[6]. The activated G-proteins are later desensitized by the intrinsic GTPase activity of the Gα subunit, followed by re-association with the Gβγ complex. Therefore, the guanine nucleotide state of the Gα subunit plays a critical role in controlling G-protein signaling [2]. In fungi, G-proteins are involved in the regulation of a variety of cellular functions in vegetative growth and/or pathogenic development, such as conidiation, infection structure differentiation, and pathogenicity [7]–[9].
Regulators of G-protein signaling (RGS) proteins primarily function as GTPase-accelerating proteins (GAPs) that promote GTP hydrolysis by the Gα subunits, thereby inactivating the G-protein and rapidly switching off G protein-coupled signaling pathways [10], [11]. All RGS proteins contain a conserved domain of ∼120 amino acids that are required for activity and function as key negative regulators of G-protein signaling pathways [12]–[14]. The budding yeast Saccharomyces cerevisiae contains four RGS and RGS-like proteins: Sst2, Rgs2, Rax1, and Mdm1. The archetypical RGS protein Sst2 possesses two N-terminal DEP (Disheveled, EGL-10, Pleckstrin) homology domains and a C-terminal RGS domain, Rgs2 has an N-terminal RGS domain, Rax1 has an N-terminal RGS domain and three C-terminal trans-membrane motifs, and Mdm1 contains an N-terminal PXA and a C-terminal PX domain in addition to an RGS domain [15].
The ascomycete Magnaporthe oryzae is pathogenic to important crops such as rice, barley, wheat, and millet. Rice blast, caused by this heterothallic haploid fungus, is one of the most severe fungal diseases of rice throughout the world [16]. Genetic studies of this important pathogen have advanced dramatically in the past decade, and thus it is an excellent model system for investigating plant–pathogen interactions. M. oryzae infects rice plants in a manner typical of many other foliar pathogens. Germ tubes produced from conidia attached to leaf surfaces differentiate into specialized infection structures called appressoria. The enormous turgor pressure generated in appressoria by the accumulation of high concentrations of glycerol is used to penetrate the underlying plant surface [17]. Mutants blocked at appressorium formation or appressorial turgor generation fail to infect healthy rice plants [18]. After penetration, infection hyphae grow in and between plant cells, and eventually result in lesion formation on the plant. Thousands of conidia are produced on the lesions and then released to initiate a new disease cycle on new plant tissues within 3–5 days. Initiation of appressorium formation in M. oryzae was shown to require G-protein and cAMP signaling, because loss of Gα MoMagB and adenylyl cyclase MoMac1 leads to failure in appressorium formation [19], [20]. A MAP kinase cascade has also been identified as an essential signaling pathway involved in appressorium formation during pathogenic development [21]–[23]. M. oryzae contains three distinct Gα proteins (MoMagA, MoMagB, and MoMagC), two Gβ subunits (MoMgb1 and MoMgb2), and one Gγ subunit [19], [24], [25]. Previous studies revealed that a constitutively active allele, MoMagBG42R, and MoMgb1 have affected G-protein signaling in vegetative growth, sexual reproduction, and pathogenicity in M. oryzae [24], [26]. The ΔMomgb1 mutant also has a defect in appressoria formation, whereas increased MoMgb1 levels promote precocious appressoria formation [24]. Moreover, expression of a dominant active allele of MoMAGB caused appressoria to form on non-inductive surfaces, while exogenous cAMP can activate appressorium formation in a ΔMomagB mutant [19], [26], [27], indicating that MoMagB may sense surface cues and stimulate cAMP synthesis. The regulator of G protein signaling Rgs1, which interacts with all three Gα subunit, was shown to negatively regulate G-protein signaling. Deletion of MoRGS1 leads to a significant increase in intracellular cAMP levels and conidiation, and ΔMorgs1 mutants also form appressoria on non-inductive hydrophilic surfaces [28]. These observations suggest that G-protein signaling and its regulators play important roles in activating the downstream cAMP pathway and regulating vegetative growth and pathogenic development. Further characterization of G-protein regulators will be helpful in better understanding the role of G-protein-mediated signaling in the regulation of early events during plant infection by the rice blast fungus.
Here, we systematically characterized all eight RGS proteins (MoRgs1–8) in M. oryzae. We found MoRgs1 has a role in regulating cell wall integrity and surface hydrophobicity, in addition to a role in mycelia growth, conidiation, sexual reproduction, and pathogenicity as previously reported [28]. All RGS proteins were involved to certain degree in the regulation of intracellular cAMP levels. Other RGS proteins, MoRgs4 in particular, also exhibit various degree of roles in conidiation, vegetative growth, asexual and sexual development, appressorium formation, and pathogenicity.
Results
M. oryzae contains eight genes encoding RGS and RGS-like proteins
MoRgs1 was first identified as a negative regulator of the G-protein signaling pathway during the important developmental events such as conidial and appressorium formation in M. oryzae [28]. To obtain a comprehensive understanding of RGS protein functions, seven additional genes encoding RGS and RGS-like proteins were identified and their biological functions characterized.
S. cerevisiae Sst2, Rgs2, Rax1, and Mdm1 RGS protein sequences were used to search the M. oryzae genome database using blastp (http://www.broadinstitute.org/annotation/genome/magnaporthe_grisea-/MultiHome.html), and each yielded a single homolog named MoRgs1 (MGG_14517.6), MoRgs2 (MGG_03146.6), MoRgs3 (MGG_03726.6), and MoRgs4 (MGG_00990.6), respectively. As expected, MoRgs1 remains as the same as previously described [28], whereas MoRgs2, MoRgs3, and MoRgs4 share high similarities to S. cerevisiae Rgs2, Rax1, and Mdm1 in the domain architectures with the amino acid sequence identity within the RGS domain being 26%, 26%, and 19% respectively (Figure 1A). In addition to these proteins, further search led to the identification of MoRgs5 (MGG_08735.6), MoRgs6 (MGG_09618.6), MoRgs7 (MGG_11693.6), and MoRgs8 (MGG_13926.6). Amino acid sequence similarities between each homolog are shown through phylogenetic analysis (Figure 1B). MoRgs5 contains an N-terminal RGS domain and a C-terminal PAS and PAC domain, while MoRgs6 possesses an N-terminal RGS domain and three C-terminal transmembrane domains. MoRgs7 and MoRgs8 appear unique it that they contain multiple transmembrane domains (seven) N-terminus of the RGS domain (Figure 1C).

Expression patterns reveal potential roles of RGS proteins in conidiogenesis, appressorium development, and infectious growth
To gain insight into the possible functions of these RGS and RGS-like proteins, we examined the gene transcription profiles during conidial stages by quantitative real-time PCR (qRT-PCR). Compared to the mycelium stage, the transcription of all MoRGS genes was upregulated; expression levels of MoRGS1 (>50-fold), MoRGS2 (>48-fold), MoRGS3 (> 44-fold), MoRGS6 (> 82-fold), MoRGS7 (> 88-fold), and were much higher than those of MoRGS4 (> four-fold), MoRGS5 (> seven-fold), and MoRGS8 (> six-fold). In the appressorium stage, MoRGS1 to MoRGS6 genes also showed high transcription levels than those during the mycelial stage. Besides MoRGS2 that three-fold increase, the other five MoRGS genes showed significant increases in transcription (MoRGS1, > 59-fold; MoRGS3, > 19-fold; MoRGS4, > 31-fold; MoRGS5, > 11-fold; MoRGS6, > 87-fold) (Table 1). During the infection stage, all eight RGS genes also showed increased expression levels; however, only the increases in MoRGS1, MoRGS3, and MoRGS7 were significant (MoRGS1 > 12-fold; MoRGS3 > 10-fold; MoRGS3 > 17-fold), in comparison to MoRGS2 (> three-fold), MoRGS4 (> 1.9-fold), MoRGS5 (> two-fold), MoRGS6 (> four-fold), and MoRGS7 (> two-fold) (Table 1). These results suggest that RGS proteins in M. oryzae likely play various roles in conidiogenesis, appressorium formation, and infection of the host plant.

Opposed roles of MoRgs1 and MoRgs4, and MoRgs2 and MoRgs3 in asexual reproduction
Mutant strains specific to each RGS gene were generated and verified (Figure S1A–D). Since G-protein and the cAMP pathway are important in conidium formation and loss of MoRgs1 and phosphodiesterase MoPdeH led to enhanced conidiation [28], [29], the role of the additional RGS proteins in conidiogenesis was examined. Under normal conditions, only ΔMorgs4 and ΔMorgs6 mutant strains exhibited darkened colony with less dense hyphal mat and only ΔMorgs1 exhibited progressive autolysis (Figure 2A). Microscopic observations indicated that ΔMorgs1 and ΔMorgs4 produced fewer conidiophores and conidia than the wild type strain. In contrast, ΔMorgs2 and ΔMorgs3 showed enhanced conidiophore and conidium formation, and ΔMorgs5, ΔMorgs6, ΔMorgs7 and ΔMorgs8 exhibited no observable changes in the production of conidiophores and conidia (Figure 2B). To validate these findings, the numbers of conidia produced from each plate were quantified. Consistently, the conidia number of ΔMorgs1 and ΔMorgs4 was reduced to 0.64-fold and 0.038-fold that of the wild type, while that of ΔMorgs2 and ΔMorgs3 was increased 1.59-fold and 1.55-fold, and ΔMorgs5, ΔMorgs6, ΔMorgs7 and ΔMorgs8 strains produced nearly the same number of conidia, 1.02-fold and 1.08-fold, as the wild type, respectively (Table 2). Our finding suggests differentiated roles in conidiophore development and conidia formation by various RGS proteins in M. oryzae: MoRgs1 and MoRgs4 have positive roles, while MoRgs2 and MoRgs3 have negative effect on the regulation of these processes.


To further evaluate the role of MoRgs1 and MoRgs4 in conidiation, a ΔMorgs1 ΔMorgs4 double mutant strain was generated and characterized (Figure S2). The mutant showed even more pronounced defect in vegetative growth with almost no conidia or conidiophores found (Figure 2C and 2D), indicating that MoRgs1 and MoRgs4 function on different targets in conidiogenesis.
Effects of MoRgs2-8 on appressorium formation
In M. oryzae, physical cues of an inductive surface, such as hardness and hydrophobicity, are required for appressorium formation [28]. However, appressorium can be induced on non-inductive surfaces in the presence of exogenous cAMP or inhibitors of cAMP phosphodiesterase [30]. Since MoRgs1 regulates cAMP levels and the ΔMorgs1 mutant formed normal appressoria on non-inductive surfaces [28], we studied the functions of other RGS and RGS-like proteins in appressorium formation. No appressoria formation was observed in mutant strains of ΔMorgs2, ΔMorgs3, ΔMorgs4, ΔMorgs5, ΔMorgs6, ΔMorgs7, and ΔMorgs8 on non-inductive surfaces (Figure 3). On inductive surfaces, all of the mutant strains, except ΔMorgs5 and ΔMorgs8, often produced two appressoria, either on branched germ tubes or on two germ tubes that emerged from one conidial cell after extended incubation of over 24 hours (Figure 3). The percentage of conidia forming two appressoria on germ tubes emerging from a single conidial cell was as high as 15% in these mutants. The appressoria formed on the secondary branching germ tubes were usually smaller than those formed on the primary germ tubes (Figure 3). These results indicate that, apart from MoRgs5 and MoRgs8, MoRgs2, MoRgs3, MoRgs4, MoRgs6, and MoRgs7 may also be involved in germ tube growth and appressoria formation.

MoRgs1 and MoRgs4 are indispensable for sexual reproduction
A previous study revealed that G-protein signaling is involved in sexual development in M. oryzae [26]. To determine whether additional proteins play any roles in mating, the ΔMorgs mutant and the wild-type strains (Guy11, MAT1-2) were crossed to a standard tester strain, TH3 (MAT1-1). After 3 weeks, numerous perithecia were observed at the junctions of the cross between the wild-type strain and TH3, complement transformants (ΔMorgs1/MoRGS1 and ΔMorgs4/MoRGS4)/TH3, but no or only a few perithecia were found for crosses between ΔMorgs1 and TH3, and ΔMorgs4 and TH3 (Figure 4, upper panel). No ascus was produced for ΔMorgs1 x TH3 and very few asci for ΔMorgs4 x TH3 (Figure 4, middle panel). These results suggest that MoRgs1 and MoRgs4 have positive roles in mating.

MoRgs1 is essential for the maintenance of cell wall integrity
In our most recent study, deletion of the MoPDEH gene encoding phosphodiesterase resulted in a cell wall integrity defect, and the ΔMopdeH mutant also underwent progressive autolysis of mycelia after incubation on CM agar plates for over 14 days [31]. This is similar to autolysis occurred in mutant strains of ΔMomps1 and ΔMomck1 mutants [32], [33]. MoMps1 and MoMck1 are homologs of S. cerevisiae Slt2 and Bck1 proteins that are involved in cell wall integrity. Because RGS proteins are generally negative regulators of G-protein signaling, deletion of RGS genes may activate downstream cAMP signals, as occurred in the ΔMopdeH mutant [31]. We tested all ΔMorgs mutants on CM agar plates for autolysis. Only the mycelia of the ΔMorgs1 mutant underwent progressive autolysis after incubation for 14 days, similar to the ΔMopdeH mutant, and none of the mutant strains showed any autolysis (Figure 2A). Moreover, the autolysis phenotype of the ΔMorgs1 mutant was suppressed by addition of 1 M sorbitol to the culture medium (Figure 2A), as also found with the ΔMopdeH and ΔMomps1 mutants [32], [33]. These results suggest that MoRgs1 plays an important role in the maintenance of cell wall integrity.
MoRgs1 and MoRgs4 are required for surface hydrophobicity
Disruption of several hydrophobin genes of M. oryzae, including MoMPG1 and MoMHP1, and MoPDEH resulted in a water - or detergent-soaked, easily wettable phenotype [34], [35], [36], [37], [38], [39], [40]. To determine whether any RGS proteins are involved in surface hydrophobicity, all ΔMorgs mutant strains were tested with water and detergent solutions. Compared with wild type and complement transformants (ΔMorgs1/MoRGS1 and ΔMorgs4/MoRGS4), none showed an easily wettable phenotype when incubated with water droplets (10 µl) after several hours. However, aerial hyphae of ΔMorgs1 and ΔMorgs4 mutants that were grown on CM agar were more readily wettable with a solution containing both 0.02% SDS and 5 mM EDTA within 5 min (Figure 5). This is similar to the ΔMomhp1 and ΔMopdeH mutants [31]. Because the expression levels of MoMPG1 and MoMHP1 were altered in the ΔMopdeH mutant [31], we speculate that the surface hydrophobicity defect may also be related to MoMpg1 and MoMhp1. To test this hypothesis, we examined the expression levels of MoMPG1 and MoMHP1. Like the ΔMopdeH mutant, the MoMPG1 expression level showed a significant decrease (> 1000-fold) in the ΔMorgs1 and ΔMorgs4 mutants, and a relatively small decrease in the ΔMorgs2 mutant (> 30-fold). In contrast, MoMHP1 expression increased to different extents in all of the mutants: the fold increase for ΔMorgs1, ΔMorgs2, ΔMorgs3, ΔMorgs4, ΔMorgs5, ΔMorgs6, ΔMorgs7, and ΔMorgs8 was two, 10, 30, three, three, six, two, and three-fold, respectively (Figure 5). These results indicate that MoRgs1 and MoRgs4 play a role in regulating surface hydrophobicity, likely through regulation of the MoMPG1 expression levels.

RGS proteins regulate MoPTH11 expression
The pathogenicity factor MoPth11 was reported to be involved in the cAMP pathway, as its transcription is regulated by exogenous cAMP [41], [42]. Disruption of MoPDEH also affected the expression of MoPTH11 during plant infection [31]. The expression of MoPTH11 was thus assessed and found to be down-regulated in all of the mutants. The transcription of MoPTH11 decreased more than 30-, 17-, 20-, and 20-fold, in ΔMorgs1, ΔMorgs2, ΔMorgs4, and ΔMorgs8 mutants, respectively. However, in the mutants ΔMorgs3, ΔMorgs5, ΔMorgs6, and ΔMorgs7, the expression of MoPth11 was not obviously changed (Figure 6). These results indicated that the RGS proteins have differentiated roles in the transcription of PTH11 in M. oryzae.

MoRgs4 affects extracellular laccase and peroxidase activities
To determine whether any RGS proteins are involved in the regulation of laccase activity, a pathogenicity factor of certain fungi [43]-[47], we tested the mutant strains on CM agar and liquid medium supplemented with 0.2 mM 2, 2′-azino-di-3-ethylbenzthiazoline-6-sulfonate (ABTS). In each case, decreases in laccase activity were only seen in the ΔMorgs4 mutant, with a less-oxidized dark purple stain around colonies of the mutant and a lower level of laccase activity in the culture filtrate compared with the wild-type strain (Figure 7A and 7B). Consistent with these observations, the expression levels of two extracellular laccase genes, MGG11608.6 and MGG13464.6, were also significantly down in ΔMorgs4 mutants (Figure 7C).

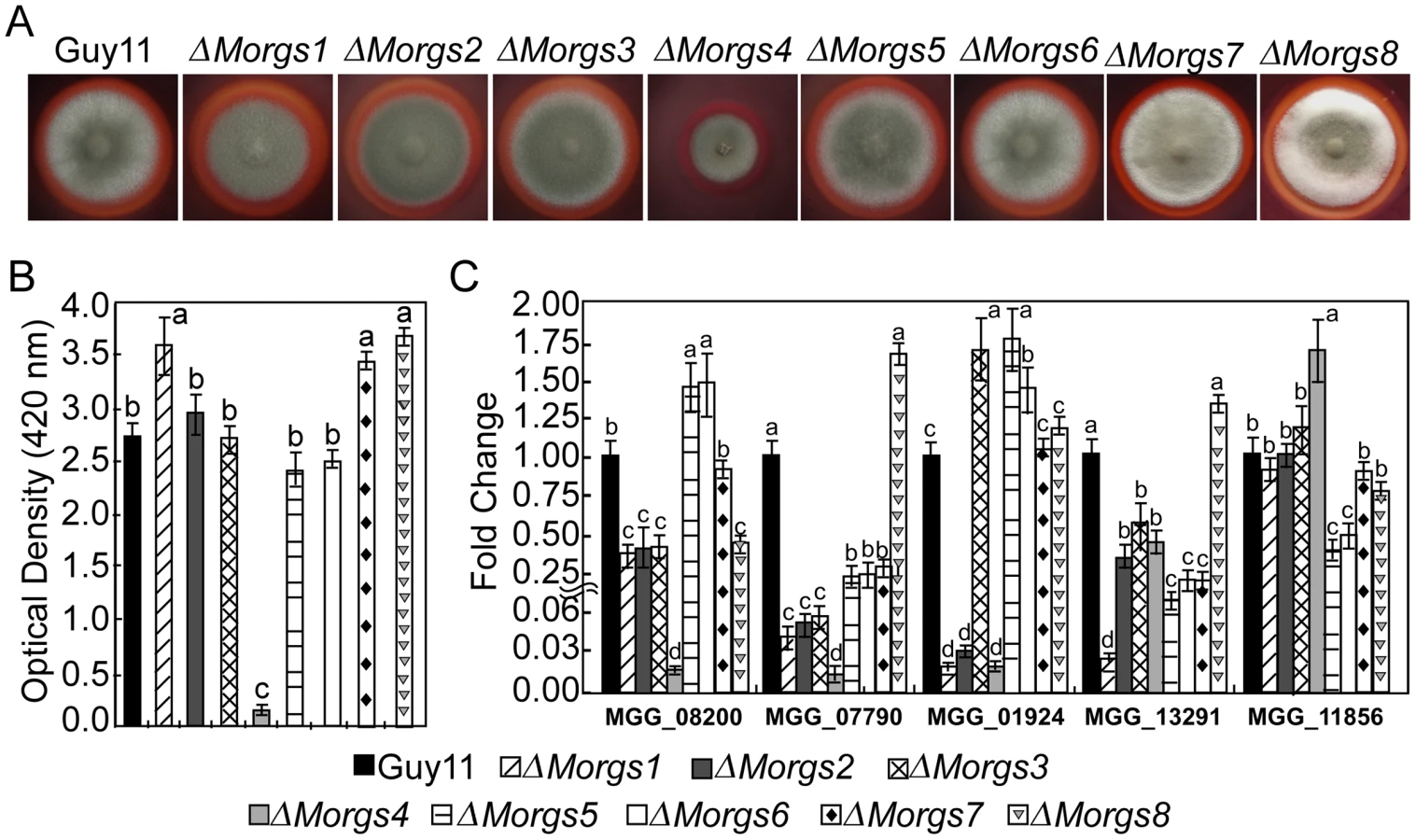
The Congo red degradation reaction is catalyzed by peroxidase, which requires H2O2 as a limiting substrate [48], [49], [50]. Discolored halos were observed beyond the wild-type colony margins when cultured on CM agar plates with Congo red, but there was no color change with the ΔMorgs4 mutant, implying that MoRgs4 is involved in peroxidase activity (Figure 8A). Enzyme activity assays using ABTS as substrate revealed that the ΔMorgs4 mutant almost lost its peroxidase activity in the extracellular culture filtrate (Figure 8B). We further examined the transcriptional level of five peroxidase-encoding genes that possess a signal peptide. The expression levels of MGG08200.6, MGG07790.6, MGG_01924.6, and MGG_13291.6 were dramatically downregulated in the ΔMorgs4 mutant and, in contrast, only MGG_11856.6 was upregulated in this mutant. A lesser degree of downregulation was observed for the other ΔMorgs mutants (Figure 8C). These data suggested that RGS proteins might be all involved in the regulation of extracellular peroxidases with MoRgs4 playing a more prominent role.
MoRgs1, MoRgs3, MoRgs4, and MoRgs7 are required for full virulence
According to the expression profiles (Table 1), the expression of RGS genes, especially MoRGS1, MoRGS3 and MoRGS7, was significant altered during infectious growth in planta, suggesting their potential roles in pathogenicity. To further test virulence involvement of these RGS proteins, susceptible rice seedlings of CO-39 were sprayed with conidia of various ΔMorgs mutants. Very few lesions were found up to 7 days post-inoculation with ΔMorgs1, ΔMorgs3, ΔMorgs4, and ΔMorgs7 mutants. In contrast, rice seedlings sprayed with ΔMorgs2, ΔMorgs5, ΔMorgs6, and ΔMorgs8 mutants under the same conditions developed numerous typical rice blast lesions similar to the wild type strain (Figure 9A). Since the ΔMorgs1, ΔMorgs3, ΔMorgs4, and ΔMorgs7 mutants exhibited normal appressoria formation, we examined the ability of the appressoria in penetration of the onion epidermal cells. Interestingly, only ΔMorgs3 and ΔMorgs7 showed decreased penetration efficiency (61% and 21%) compared to the wild type (85%), while no change was found for the other ΔMorgs mutants (Table 2). However, the majority of the appressoria in the ΔMorgs2, ΔMorgs5, ΔMorgs6, and ΔMorgs8 mutants formed invasive hyphae, while only a few limited infectious hyphae developed from appressoria of ΔMorgs1, ΔMorgs3, ΔMorgs4, and ΔMorgs7 mutants (data not shown). To further validate this observation, we repeated the test with rice sheath cells and found that the appressoria of the ΔMorgs2, ΔMorgs5, ΔMorgs6, and ΔMorgs8 mutants as well as the wild type strain, produced branching infectious hyphae, but the appressoria of the ΔMorgs1, ΔMorgs3, ΔMorgs4, and ΔMorgs7 mutants produced only limited infectious hyphae (Figure 9B). These results indicate that MoRgs1, MoRgs3, MoRgs4, and MoRgs7 have significantly reduced virulence resulting from the defect in infectious hyphal growth.

Moreover, we examined the collective effect of MoRgs1 and MoRgs4 on pathogenicity on detached barley leaves. Consistently, ΔMorgs1 and ΔMorgs4 were less virulent than the wild type strain Guy11 and the ΔMorgs1ΔMorgs4 double mutant strain lost all pathogenicity (Figure 9C). Further observation indicated that the ΔMorgs1ΔMorgs4 was unable to form appressorium on induction surfaces (Figure 9D).
Functions of M. oryzae RGS proteins in cAMP and G protein signaling
MoRgs1 plays an important role in regulation of the intracellular cAMP level in M. oryzae [28]. To determine whether other RGS and RGS-like proteins are also involved in this process, we measured intracellular cAMP levels of the mutants in the hyphal stage and compared with that of the wild-type and the ΔMomac1 mutant strains. The results indicated that all ΔMorgs mutant strains accumulate somewhat higher levels of cAMP than the wild-type strain. Compared with wild-type, ΔMorgs1 showed a ∼3.7-fold higher intracellular cAMP level, which is consistent with the earlier study [28]. Surprisingly, an increase of five-fold was found in the ΔMorgs2 mutant while the ΔMorgs3-8 mutants accumulated 2.8-, 2.8-, 2.2-, 2.1-, 3-, and 3-fold higher levels of cAMP, respectively (Figure 10). While the mechanism remains unclear, particularly for ΔMorgs4 and ΔMorgs5 mutant strains, these results suggest that RGS proteins have important roles in regulating intracellular cAMP levels.

The RGS domain of RGS proteins has a high affinity to Gα and the binding specificity between RGS and Gα proteins often determine signal specificity and amplitude. In S. cerevisiae, Sst2 functions as a negative regulator of pheromones and mating by interacting with Gα Gpa1 [12] and Rgs2 down-regulates glucose activation of the cAMP pathway through direct inhibition of Gpa2 [51]. In a study by Chasse et al., Sst2, Rgs2, Rax1, and Mdm1 were all found to bind Gpa1 and affect Gpa1 signaling, although Sst2 still remained the most regulatory role in Gpa1 signaling and mating [15]. M. oryzae MoRgs1 was found to interact with MoMagA for pathogenicity and MoMagB for conidiation [28]. To find out whether other RGS proteins also function similarly by binding to all or specific Gα proteins, a yeast two-hybrid (Y2H) assay was conducted. In this assay, MoRgs2, MoRgs5, MoRgs7 and MoRgs8 were found to interact with MoMagB, while MoRgs7 only interacted with MoMagA. Surprisingly, all RGS and RGS-like proteins interacted with MoMagC (Figure 11A and 11B). MoRgs1 failed to interact with MoMagA in contrast to previously reported may due to the different experimental conditions utilized [28].

To further investigate the interaction between MoMagB and MoRgs2, a ΔMomagBΔMorgs2 double mutant strain was generated and analyzed. In addition, two RGS-insensitive mutant alleles, MoMAGBG183S and MoMAGBQ204L allele were also introduced respectively into the ΔMomagB mutant. Consistent with a role for MoRgs2 in conidiation that is mediated through MoMagB, no conidia was found in the ΔMomagBΔMorgs2 mutant, whereas a significant increase in conidiation was found for ΔMorgs2 expressing MoMABG183S and MoMAGBQ204L alleles (Figure 12).

These results indicated that MoRgs2 functions upstream of MoMagB in conidiation. MoRgs7 regulates pathogenicity may also be mediated through MoMagB, similar to MoRgs1. The interactions between MoRgs5 and MoRgs8 and MoMagB could confer additional roles in controlling various developmental processes.
Discussion
Heterotrimeric G-proteins play crucial roles in the regulation of fungal developmental processes and pathogenicity [7]. RGS proteins function as negative regulators to enhance the intrinsic GTPase activity of Gα subunits, thereby inactivating the G-proteins and rapidly switching off the cellular response. A large number of mammalian RGS proteins have been shown to play important roles in various signaling pathways [52]. Many fungal RGS protein homologs have also been well characterized, such as Sst2 of S. cerevisiae [12], FlbA and RgsA of Aspergillus nidulans [53]–[55], CPRGS-1 of Cryphonectria parasitica [56], and Cag8 of Metarhizium anisopliae [57], as well as MoRgs1 of M. oryzae [28]. Here, we identified and characterized seven RGS proteins in addition to MoRgs1 of M. oryzae. Our findings revealed that there are as many as eight RGS and RGS-like proteins in M. oryzae, the most ever found in fungi, which play various roles in the modulation of vegetative growth, asexual/sexual development, cell wall integrity, surface hydrophobicity, appressorium formation and penetration, and pathogenicity in this pathogenic fungus (Table S2).
The M. oryzae MoRgs1 is highly homologous to A. nidulans FlbA and C. parasitica CPRGS-1. Consistent with studies of FlbA, and CPRGS-1 that positively regulate asexual development [54], [56], our data indicated that MoRgs1 positively regulates asexual sporulation. This, however, contradicted with the previous report that MoRgs1 has a negatively role in asexual development reported. Regardless, our findings for the role of MoRgs1 are mostly consistent with the previous study [28]. In M. oryzae, deletion of the gene encoding MoMagA or MoMagC has no effect on vegetative growth, appressorium formation, or pathogenicity, except that deletion of MoMAGC encoding MoMagC exhibited reduced conidiation [19]. The ΔMomagB mutant, however, exhibited significantly reduced vegetative growth, conidiation, and appressorium formation, as well as virulence [19]. A reduction in perithecium formation was observed in ΔMomagA, but not ΔMomagC, and no perithecium was found in ΔMomagB (Figure S3). The strains with the putative dominant active MoMAGBG42R allele formed appressoria on both hydrophobic and hydrophilic surfaces [26]. This phenotype was also observed in the transformant carrying multiple copies of MoMGB1 [24] and in a phosphodiesterase gene mutant ΔMopdeH [31], similar to results observed in the ΔMorgs1 mutant, suggesting that expression of MoMAGBG42R and multiple copies of MoMGB1, inactivation of MoPDEH, and deletion of MoRGS1 all enhance cAMP signaling. Because of this, it is feasible to hypothesize that MoRgs1 has a negative effect on Gα MoMagB signaling. The MoMAGBG42R mutation also reduced conidiation and virulence, failed to form asci, and caused autolysis of aged colonies [26], and the same phenotypes appeared in the ΔMorgs1 mutant, suggesting that MoRgs1 negatively regulates MoMagB. Surprisingly, MoRgs1 and MoMagB failed to interact with each other in our hand.
Instead, interactions between MoRgs7 and MoMagA, MoRgs2, MoRgs5, MoRgs7, MoRgs8 and MoMagB, and MoRgs1-8 and MoMagC were found. These interactions could suggest functional significance (differentiated interactions with MoMagA and MoMagB) or not (interactions with MoMagC). Indeed, MoRgs1 and MoRgs4 likely positively regulate MoMagB in conidiation, whereas MoRgs2 and MoRgs3 negatively regulate MoMagB and or MoMagC in this process. For sexual reproduction, MoRgs4 may be involved in a similar mechanism to MoRgs1, but extra and different regulators must exist in the regulation of MoRgs1 and MoRgs4, since the ΔMorgs4 mutant produced few asci while the ΔMorgs1 mutant produced no asci. Moreover, MoRgs2, MoRgs5, MoRgs7 and MoRgs8 were found to interact with MoMagB. While no phenotypic changes exhibited by MoRgs5, the ΔMorgs2 mutant did show increased conidiation, suggesting that MoRgs2 may negatively regulate MoMagB in conidiogenesis and pathogenesis. Since MoRgs4 and MoRgs5 are structurally more similar to S. cerevisiae Mdm1 whose role in G protein was not established, their roles in M. oryzae may be established independent of G protein signaling.
Interestingly, with the exception of ΔMorgs5 and ΔMorgs8, most ΔMorgs mutants often formed multiple appressoria on hydrophobic surfaces, which were also observed in the site-directed mutation transformant MoMGB1D41N, as the aspartic acid residue at 41 is known to be involved in the interaction between MoMgb1 and MgSte20 [24]. This may indicate that RGS proteins could play a role in activating the D41 of MoMgb1 or have cross-talk between MoSte20 (MgSte20) signaling for germ tube growth and correct regulation of appressorium formation.
The fungal cell wall plays important roles during cell division, growth, and morphogenesis, and in mediating all exchanges between the cell and its environment [58], [59]. In pathogenic fungi, the ability to maintain cell wall integrity is critical to the establishment of disease in the host [33]. Several cell wall integrity-associated genes such as MoMps1 and MoMck1 have been characterized in M. oryzae. MoMps1 and MoMck1 have been described as essential for cell wall integrity and pathogenicity [32], [33]. In our latest studies, a constitutive activating cAMP pathway mutant ΔMopdeH also showed an autolysis phenotype like that observed in the ΔMorgs1 mutant [31]. Thus, the G-protein/cAMP signaling pathway may have cross-talk with the MAPK pathway in regulating cell wall integrity. Alternatively, like the ΔMopdeH mutant, the cell wall integrity defect in ΔMorgs1 could be due to the high intracellular cAMP level in the mutant. However, based on our results, there should be more regulators involved in regulation of cell wall integrity, because other ΔMorgs mutants besides ΔMorgs1 also have high intracellular cAMP levels but do not exhibit the autolysis phenotype.
Most hydrophobins confer surface hydrophobicity on fungi forming a spore rodlet layer. Deletion of several hydrophobin genes, including MoMPG1, resulted in a water - or detergent-soaked, easily wettable phenotype and these genes played important roles in multiple infection-related processes [34]–[39]. In previous studies, G-protein and cAMP signaling pathways have been reported to be involved in hydrophobin synthesis and surface hydrophobicity [31], [57]. In the insect pathogenic fungus M. anisopliae, loss of the MoRGS1 homolog gene Mocag8 reduced the transcription of a hydrophobin-encoding gene [57]. In the chestnut blight fungus C. parasitica, the RGS protein CPRGS-1 is also known to regulate hydrophobin synthesis [56]. In M. oryzae, a constitutive activate cAMP signaling mutant ΔMopdeH showed a defect in surface hydrophobicity and a low level of MoMPG1 expression [31]. These results well support the surface hydrophobicity defects of the ΔMorgs1 and ΔMorgs4 mutants. Furthermore, they can also be taken as evidence explaining the cell wall integrity defect in ΔMorgs1.
The high expression level of RGS genes at late infection stages in infected rice leaves indicates their potential role in infectious growth and virulence. Mutation of MoRGS1, MoRGS3, MoRGS4, and MoRGS7 significantly reduced virulence and produced fewer lesions than the wild-type strain, implying that appressoria formed by these mutants are probably defective in penetration. It is likely that these four genes regulate processes involved in the early stages of appressorium penetration, such as development of the penetration peg or differentiation of infectious hyphae. The reduction in pathogenicity may be due to a reduction in development at the pre-penetration stages or a defect in infectious growth of ΔMorgs mutants in host cells. According to the penetration data, MoRGS1, MoRGS3 and MoRGS7 are indeed involved in penetration and infectious growth and well support the hypothesis. However, unlike ΔMorgs1, ΔMorgs3 and MoRGS7, the reduced virulence of ΔMorgs4 mainly resulted from the defect in infectious hyphal growth, indicating the unique functions of each RGS protein in M. oryzae. RGS proteins may act as different regulators in the regulation of different targets (such as Gα subunits in pathogenesis).
In the present study, ΔMorgs4 totally lost laccase and peroxidase activity. Laccases are copper-containing oxidases found in many plants, fungi, and microorganisms. Laccase activity has been reported to be involved in virulence in some fungi [43]. In M. oryzae, several virulence attenuation mutants also show loss or reduction of laccase and peroxidase activities [44], [45], [60]. Therefore, the loss of laccase and extracellular peroxidase activity might be one aspect of the reduced virulence in the ΔMorgs4 mutant. However, ΔMorgs1 and ΔMorgs3 also had reduced pathogenicity but with normal laccase and extracellular peroxidase activity, indicating regulators other than laccases and extracellular peroxidases must exist in the G-protein signaling pathway to control infection-related processes in M. oryzae. The disparity may also indicate that the function of MoRgs4 may be more in line with that of S. cerevisiae Mdm1 and could be independent of G protein signaling.
Overall, our results indicate that different RGS proteins control unique signal transduction pathways in M. oryzae, which are involved in asexual/sexual development, appressorium differentiation, penetration, and infectious growth. It will be important and interesting to distinguish specific functions associated with each RGS proteins and link G-protein signaling to the pathogenicity of the fungus.
Materials and Methods
Strains and culture conditions
The M. oryzae Guy11 and 70–15 strains were used as wild type for transformation in this study. All strains were cultured on complete medium (CM) agar plates [31]. Liquid CM medium was used to prepare the mycelia for DNA and RNA extraction. For conidiation, strain blocks were maintained on straw decoction and corn (SDC) agar media [31] at 28°C for 7 days in the dark followed by 3 days of continuous illumination under fluorescent light.
Targeted gene deletion and complementation
Standard DNA and RNA manipulations were performed as described previously [61]. The gene-deletion mutants were generated using the standard one-step gene replacement strategy. First, two 1.0 kb of sequences flanking of targeted gene were PCR amplified with primer pairs (Table S1), then a ∼2-kb fragment containing the two flanking sequences was amplified by overlap PCR. All amplified sequences and fragments were sequenced and then ligated to flank the hygromycin resistance cassette, which was amplified with primers FL1111 & FL1112 (Table S1), into the pMD19-T vector (Takara Co. Dalian, China). The ∼3.4-kb fragments, which contain the flanking sequences and hygromycin cassette, were amplified and transformed into protoplasts of wild type Guy11. The complement fragments, which contain the entire RGS genes and their native promoter regions, were amplified by PCR with primers (Table S1) and inserted into pCB1532 (sulphonylurea resistance) or pYF11 (bleomycin resistance) to complement the mutant strains, respectively.
Vegetative growth
Small agar blocks were cut from the edge of 4-day-old cultures and placed onto CM and SDC media for culturing in the dark at 28°C. The size and morphology of the colonies were examined each day for 10 days and then photographed. The experiment was performed in triplicate.
Hyphal growth and surface hydrophobicity assay
For hyphal growth, small agar blocks were cut from the edge of 4-day-old cultures and placed onto the CM and CM adding 1 M sorbitol and cultured in the dark at 28°C for two weeks. The size and morphology of the colonies were examined every day and photographed on day 14 after incubation. For surface hydrophobicity assay, the strains were plated onto CM agar plates and incubated at 28°C for 14-day. Sterile distilled water (10 µl) was placed on the surface of cultures. In addition, wettability of aerial hyphae to solution containing both 0.02% SDS and 5 mM EDTA was also assessed as previously described [62].
Appressorium formation, cuticle penetration, and infection assays
Conidia were harvested from 10-day-old cultures, filtered through three layers of lens paper, and resuspended to a concentration of 5×104 spores per milliliter in sterile water. For appressorium formation and cuticle penetration assays, droplets (30 µl) of conidial suspension were placed on plastic cover slips (hydrophobic), Gelbond films (hydrophilic) and onion epidermal cells and incubated under humid conditions at room temperature as described previously [63]. Appressorium formation and development of invasive hyphae were examined after incubation for 24 hours. For plant infection assays, conidia were resuspended to a concentration of 5×104 spores per milliliter in a 0.2% (w/v) gelatin solution. Two-week-old seedlings of rice (Oryza sativa cv CO39) were sprayed with 5 ml of conidial suspension of each treatment. Inoculated plants were kept in a growth chamber at 25°C with 90% humidity and in the dark for the first 24 hours, followed by a 12/12 hours light/dark cycle [64]. Lesion formation was observed daily and photographed 7 days after inoculation.
For microscopic observation of penetration and infectious hyphae expansion in rice tissue, rice cultivar CO-39 were prepared as previously described [44] and inoculated with 100 µl of conidial suspension (1×104 spores per milliliter) on the inner leaf sheath cuticle cells. After 48 hours incubation under humid conditions at room temperature, the leaf sheaths were observed under a microscope. Appressorium turgor was measured by incipient cytorrhysis (cell collapse) assay using a 1–5 molar concentration of glycerol solution as described previously [65].
Mating
Plugs of ΔMorgs mutants and control strain Guy11 (MAT1-2) and the mating partner strain TH3 (MAT1-1) were point-inoculated 3 cm apart on oatmeal agar medium and incubated at 20°C under constant fluorescent light for 3 to 4 weeks. Mature perithecia were crushed to examine the asci and ascospores approximately 20 to 25 days post-inoculation.
Intracellular cAMP, laccase and peroxidase activities assays
Two-day-old liquid mycelial cultures were harvested, frozen in liquid nitrogen and lyophilized for 16 hours. Intracellular cAMP extraction was followed as previously described [28]. The cAMP levels were quantified according to the cAMP Biotrak Immuno-assay System (BD Bioscience, NJ, USA).
Laccase and peroxidase activities were measured from 2-day-old CM liquid cultures. Mycelia were removed completely by filtration and centrifugation (5,000 g at 4°C) and processed using a colorimetric determination as described previously [66].
Yeast two-hybrid assay
The bait constructs were generated by cloning MoMAGA, MoMAGB and MoMAGC full-length cDNAs into pGBKT7, respectively. The RGS cDNAs (MoRGS1, MoRGS4, MoRGS5, MoRGS6, MoRGS7 and MoRGS8: RGS domain only; MoRGS2 and MoRGS3: full-length) were cloned into pGADT7 as the prey constructs (see primers in Table S1). The resulting prey and bait constructs were confirmed by sequencing analysis and transformed in pairs into yeast strain AH109 as the description of BD library construction & screening kit (Clontech, USA). The Trp+ and Leu+ transformants were isolated and assayed for growth on SD-Trp-Leu-His-Ade medium and the expression of LacZ reporter gene following the instructions provided by Clontech. Yeast stains for positive and negative controls were from the Kit.
Co-immunoprecipitation (co-IP) and western blot analysis
The MoMAGB-3xFLAG and MoRGS2-GFP constructs were generated with the yeast gap repair approach [23], [67] and confirmed by sequencing analysis. The resulting fusion constructs were co-transformed into protoplasts of 70-15. Transformants expressing the MoMAGB-3xFLAG and MoRGS2-GFP constructs were identified by PCR and confirmed by western blot analysis with an anti-FLAG antibody (Sigma-Aldrich, USA). For co-IP assays, total proteins were isolated from vegetative hyphae as described [23] and incubated with anti-FLAG M2 beads (Sigma-Aldrich). Western blots of proteins eluted from the M2 beads were detected with the anti-GFP [23], anti-FLAG and anti-actin (Sigma-Aldrich) antibodies with the ECL Supersignal System (Pierce, USA).
Construction of the MoMAGBG183S and MoMAGBQ204L alleles
PCR products containing the native promoter of MoMAGB were amplified with primers FL9963/FL9965 (Table S1) and co-transformed with fragments amplified with primers FL9966/FL9964 (Table S1) into the yeast strain XK1-25 with XhoI digested vector pYF11 that contains the bleomycin-resistant gene and the GFP gene [23]. Plasmid pYF11::MoMAGBG183S was rescued from the resulting Trp+ yeast transformants. The same strategy was used to generate the pYF11::MoMAGBQ204L vector (PCR products amplified with primers FL9963/FL9967 and FL9968/FL9964, respectively, Table S1). Protoplasts of the Guy11 and ΔMorgs2 mutant were transformed with pYF11::MoMAGBG183S or pYF11::MoMAGBQ204L.
Supporting Information
{kind=link}
{kind=link}
Zdroje
1. MalbonCC 2005 G proteins in development. Nat Rev Mol Cell Bio 6 689 701
2. DohlmanHGThornerJW 2001 Regulation of G protein-initiated signal transduction in yeast: Paradigms and principles. Annu Rev Biochem 70 703 754
3. ClaphamDENeerEJ 1997 G protein beta gamma subunits. Annu Rev Pharmacol Toxicol 37 167 203
4. HammHE 1998 The many faces of G protein signaling. J Biol Chem 273 669 672
5. NeerEJ 1995 Heterotrimeric G-proteins - organizers of transmembrane signals. Cell 80 249 257
6. SimonMIStrathmannMPGautamN 1991 Diversity of G-proteins in signal transduction. Science 252 802 808
7. BolkerM 1998 Sex and crime: Heterotrimeric G proteins in fungal mating and pathogenesis. Fungal Genet Biol 25 143 156
8. LengelerKBDavidsonRCD'SouzaCHarashimaTShenWC 2000 Signal transduction cascades regulating fungal development and virulence. Microbiol Mol Biol Rev 64 746 785
9. YuJH 2006 Heterotrimeric G protein signaling and RGSs in Aspergillus nidulans. J Microbiol 44 145 154
10. SiderovskiDPWillardFS 2005 The GAPs, GEFs, and GDIs of heterotrimeric G-protein alpha subunits. Int J Biol Sci 1 51 66
11. De VriesLZhengBFischerTElenkoEFarquharMG 2000 The regulator of G protein signaling family. Annu Rev Pharmacol Toxicol 40 235 271
12. DohlmanHGSongJPMaDRCourchesneWEThornerJ 1996 Sst2, a negative regulator of pheromone signaling in the yeast Saccharomyces cerevisiae: Expression, localization, and genetic interaction and physical association with Gpa1 (the G-protein alpha subunit). Mol Cell Biol 16 5194 5209
13. KoelleMRHorvitzHR 1996 EGL-10 regulates G protein signaling in the C-elegans nervous system and shares a conserved domain with many mammalian proteins. Cell 84 115 125
14. SiderovskiDPHesselAChungSMakTWTyersM 1996 A new family of regulators of G-protein-coupled receptors? Curr Biol 6 211 212
15. ChasseSAFlanaryPParnellSCHaoNChaJY 2006 Genome-scale analysis reveals Sst2 as the principal regulator of mating pheromone signaling in the yeast Saccharomyces cerevisiae. Eukaryot Cell 5 330 346
16. ValentB 1990 Rice blast as a model system for plant pathology. Phytopathology 80 33 36
17. de JongJCMcCormackBJSmirnoffNTalbotNJ 1997 Glycerol generates turgor in rice blast. Nature 389 244 245
18. ValentBFarralLChumleyFG 1991 Magnaporthe grisea genes for pathogenicity and virulence identified through a series of backcrosses. Genetics 127 87 101
19. LiuSHDeanRA 1997 G protein alpha subunit genes control growth, development, and pathogenicity of Magnaporthe grisea. Mol Plant Microbe Interact 10 1075 1086
20. ChoiWBDeanRA 1997 The adenylate cyclase gene MAC1 of Magnaporthe grisea controls appressorium formation and other aspects of growth and development. Plant Cell 9 1973 1983
21. XuJRHamerJE 1996 MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Genes Dev 10 2696 2706
22. XuJR 2000 MAP kinases in fungal pathogens. Fungal Genet Biol 31 137 152
23. BrunoKSTenjoFLiLHamerJEXuJR 2004 Cellular localization and role of kinase activity of PMK1 in Magnaporthe grisea. Eukaryot Cell 3 1525 1532
24. NishimuraMParkGXuJR 2003 The G-beta subunit MGB1 is involved in regulating multiple steps of infection-related morphogenesis in Magnaporthe grisea. Mol Microbiol 50 231 243
25. DeanRATalbotNJEbboleDJFarmanMLMitchellTK 2005 The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 434 980 986
26. FangEGCDeanRA 2000 Site-directed mutagenesis of the MAGB gene affects growth and development in Magnaporthe grisea. Mol Plant Microbe Interact 13 1214 1227
27. EbboleDJ 2007 Magnaporthe as a model for understanding host-pathogen interactions. Annu Rev Phytopathol 45 437 456
28. LiuHSureshAWillardFSSiderovskiDPLuS 2007 Rgs1 regulates multiple G alpha subunits in Magnaporthe pathogenesis, asexual growth and thigmotropism. EMBO J 26 690 700
29. RamanujamRNaqviNI 2010 PdeH, a high-affinity cAMP phosphodiesterase, is a key regulator of asexual and pathogenic differentiation in Magnaporthe oryzae. PLoS Pathog 6 e1000897
30. LeeYHDeanRA 1993 cAMP regulates infection structure formation in the plant pathogenic fungus Magnaporthe grisea. Plant Cell 5 693 700
31. ZhangHFLiuKYZhangXTangWWangJS 2010 Two phosphodiesterase genes, PDEL and PDEH, regulate development and pathogenicity by modulating intracellular cyclic AMP levels in Magnaporthe oryzae. PLoS One 6 e17241
32. XuJRStaigerCJHamerJE 1998 Inactivation of the mitogen-activated protein kinase Mps1 from the rice blast fungus prevents penetration of host cells but allows activation of plant defense responses. Proc Natl Acad Sci U S A 95 12713 12718
33. JeonJGohJYooSChiMHChoiJ 2008 A putative MAP kinase kinase kinase, MCK1, is required for cell wall integrity and pathogenicity of the rice blast fungus, Magnaporthe oryzae. Mol Plant Microbe Interact 21 525 534
34. StringerMADeanRASewallTCTimberlakeWE 1991 Rodletless, a new Aspergillus developmental mutant induced by directed gene inactivation. Genes Dev 5 1161 1171
35. BellpedersenDDunlapJCLorosJJ 1992 The Neurospora circadian clock-controlled gene, Ccg-2, is allelic to Eas and encodes a fungal hydrophobin required for formation of the conidial rodlet layer. Genes Dev 6 2382 2394
36. LauterFRRussoVEAYanofskyC 1992 Developmental and light regulation of Eas, the structural gene for the rodlet protein of Neurospora. Genes Dev 6 2373 2381
37. TalbotNJEbboleDJHamerJE 1993 Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 5 1575 1590
38. vanWetterMASchurenFHJSchuursTAWesselsJGH 1996 Targeted mutation of the SC3 hydrophobin gene of Schizophyllum commune affects formation of aerial hyphae. FEMS Microbiol Lett 140 265 269
39. SpanuP 1998 Deletion of HCf-1, a hydrophobin gene of Cladosporium fulvum, does not affect pathogenicity in tomato. Physiol Mol Plant Pathol 52 323 334
40. KimSAhnIPRhoHSLeeYH 2005 MHP1, a Magnaporthe grisea hydrophobin gene, is required for fungal development and plant colonization. Mol Microbiol 57 1224 1237
41. DeZwaanTMCarrollAMValentBSweigardJA 1999 Magnaporthe grisea Pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues. Plant Cell 11 2013 2030
42. OhYDonofrioNPanHQCoughlanSBrownDE 2008 Transcriptome analysis reveals new insight into appressorium formation and function in the rice blast fungus Magnaporthe oryzae. Genome Biol 9 R85
43. BarnunNLevATHarelEMayerAM 1988 Repression of laccase formation in Botrytis cinerea and its possible relation to phytopathogenicity. Phytochemistry 27 2505 2509
44. GuoMGuoWChenYDongSMZhangX 2010 The basic leucine zipper transcription factor Moatf1 mediates oxidative stress responses and is necessary for full virulence of the rice blast fungus Magnaporthe oryzae. Mol Plant Microbe Interact 23 1053 1068
45. ZhangHFLiuKYZhangXSongWWZhaoQ 2010 A two-component histidine kinase, MoSLN1, is required for cell wall integrity and pathogenicity of the rice blast fungus, Magnaporthe oryzae. Curr Genet 56 517 528
46. DouXYWangQQiZQSongWWWangW 2011 MoVam7, a conserved SNARE involved in vacuole assembly, is required for growth, endocytosis, ROS accumulation, and pathogenesis of Magnaporthe oryzae. PLoS One 6 e16439
47. GuoMChenYDuYDongYHGuoW 2011 The bZIP transcription factor MoAP1 mediates the oxidative stress response and is crucial for pathogenicity of the rice blast fungus Magnaporthe oryzae. PLoS Pathog 7 e1001302
48. CrippsCBumpusJAAustSD 1990 Biodegradation of azo and heterocyclic dyes by Phanerochaete chrysosporium. Appl Environ Microbiol 56 1114 1118
49. WooSWChoJSHurBKShinDHRyuKG 2003 Hydrogen peroxide, its measurement and effect during enzymatic decoloring of Congo red. J of Microbiol Biotechn 13 773 777
50. ChiMHParkSYKimSLeeYH 2009 A novel pathogenicity gene is required in the rice blast fungus to suppress the basal defenses of the host. PLoS Pathog 5 e1000401
51. VerseleMde WindeJHTheveleinJM 1999 A novel regulator of G protein signalling in yeast, Rgs2, downregulates glucose-activation of the cAMP pathway through direct inhibition of Gpa2. EMBO J 18 5577 5591
52. HollingerSHeplerJR 2002 Cellular regulation of RGS proteins: Modulators and integrators of G protein signaling. Pharmacol Rev 54 527 559
53. LeeBNAdamsTH 1994 Overexpression of flbA, an early regulator of Aspergillus asexual sporulation, leads to activation of brlA and premature initiation of development. Mol Microbiol 14 323 334
54. YuJHWieserJAdamsTH 1996 The Aspergillus flbA RGS domain protein antagonizes G protein signaling to block proliferation and allow development. EMBO J 15 5184 5190
55. HanKHSeoJAYuJH 2004 Regulators of G-protein signalling in Aspergillus nidulans: RgsA downregulates stress response and stimulates asexual sporulation through attenuation of GanB (G alpha) signalling. Mol Microbiol 53 529 540
56. SegersGCRegierJCNussDL 2004 Evidence for a role of the regulator of G-protein signaling protein CPRGS-1 in Galpha subunit CPG-1-mediated regulation of fungal virulence, conidiation, and hydrophobin synthesis in the chestnut blight fungus Cryphonectria parasitica. Eukaryot Cell 3 1454 1463
57. FangWPeiYBidochkaMJ 2007 A regulator of a G protein signalling (RGS) gene, cag8, from the insect-pathogenic fungus Metarhizium anisopliae is involved in conidiation, virulence and hydrophobin synthesis. Microbiol-Sgm 153 1017 1025
58. CabibERohDHSchmidtMCrottiLBVarmaA 2001 The yeast cell wall and septum as paradigms of cell growth and morphogenesis. J Bio Chem 276 19679 19682
59. AdamsDJ 2004 Fungal cell wall chitinases and glucanases. Microbiol-Sgm 150 2029 2035
60. SongWWDouXYQiZQWangQZhangX 2010 R-SNARE homolog MoSec22 is required for conidiogenesis, cell wall integrity, and pathogenesis of Magnaporthe oryzae. PLoS One 5 e13193
61. LivakKJSchmittgenTD 2001 Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25 402 408
62. StringerMATimberlakeWE 1995 Dewa encodes a fungal hydrophobin component of the Aspergillus spore wall. Mol Microbiol 16 33 44
63. ZhangHFZhaoQLiuKYZhangZGWangYC 2009 MgCRZ1, a transcription factor of Magnaporthe grisea, controls growth, development and is involved in full virulence. FEMS Microbiol Lett 293 160 169
64. ZhangLSLvRLDouXYQiZQHuaCL 2011 The function of MoGlk1 in integration of glucose and ammonium utilization in Magnaporthe oryzae. PLoS One 6 e22809
65. HowardRJFerrariMARoachDHMoneyNP 1991 Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc Natl Acad Sci U S A 88 11281 11284
66. ChiMHParkSYKimSLeeYH 2009 A novel pathogenicity gene is required in the rice blast fungus to suppress the basal defenses of the host. PLoS Pathog 5 e1000401
67. BourettTMSweigardJACzymmekKJCarrollAHowardRJ 2002 Reef coral fluorescent proteins for visualizing fungal pathogens. Fungal Genet Biol 37 211 220
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2011 Číslo 12
- Farmakovigilanční studie perorálních antivirotik indikovaných v léčbě COVID-19
- Jak souvisí postcovidový syndrom s poškozením mozku?
- Měli bychom postcovidový syndrom léčit antidepresivy?
- 10 bodů k očkování proti COVID-19: stanovisko České společnosti alergologie a klinické imunologie ČLS JEP
Nejčtenější v tomto čísle
- Controlling Viral Immuno-Inflammatory Lesions by Modulating Aryl Hydrocarbon Receptor Signaling
- Fungal Virulence and Development Is Regulated by Alternative Pre-mRNA 3′End Processing in
- Cryo Electron Tomography of Herpes Simplex Virus during Axonal Transport and Secondary Envelopment in Primary Neurons
- Epstein-Barr Virus Nuclear Antigen 3C Stabilizes Gemin3 to Block p53-mediated Apoptosis