
Interleukin-13 Promotes Susceptibility to Chlamydial Infection of the Respiratory and Genital Tracts
Chlamydiae are intracellular bacteria that commonly cause infections of the respiratory and genital tracts, which are major clinical problems. Infections are also linked to the aetiology of diseases such as asthma, emphysema and heart disease. The clinical management of infection is problematic and antibiotic resistance is emerging. Increased understanding of immune processes that are involved in both clearance and immunopathology of chlamydial infection is critical for the development of improved treatment strategies. Here, we show that IL-13 was produced in the lungs of mice rapidly after Chlamydia muridarum (Cmu) infection and promoted susceptibility to infection. Wild-type (WT) mice had increased disease severity, bacterial load and associated inflammation compared to IL-13 deficient (−/−) mice as early as 3 days post infection (p.i.). Intratracheal instillation of IL-13 enhanced bacterial load in IL-13−/ − mice. There were no differences in early IFN-g and IL-10 expression between WT and IL-13−/ − mice and depletion of CD4+ T cells did not affect infection in IL-13−/ − mice. Collectively, these data demonstrate a lack of CD4+ T cell involvement and a novel role for IL-13 in innate responses to infection. We also showed that IL-13 deficiency increased macrophage uptake of Cmu in vitro and in vivo. Moreover, the depletion of IL-13 during infection of lung epithelial cells in vitro decreased the percentage of infected cells and reduced bacterial growth. Our results suggest that enhanced IL-13 responses in the airways, such as that found in asthmatics, may promote susceptibility to chlamydial lung infection. Importantly the role of IL-13 in regulating infection was not limited to the lung as we showed that IL-13 also promoted susceptibility to Cmu genital tract infection. Collectively our findings demonstrate that innate IL-13 release promotes infection that results in enhanced inflammation and have broad implications for the treatment of chlamydial infections and IL-13-associated diseases.
Published in the journal:
. PLoS Pathog 7(5): e32767. doi:10.1371/journal.ppat.1001339
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1001339
Summary
Chlamydiae are intracellular bacteria that commonly cause infections of the respiratory and genital tracts, which are major clinical problems. Infections are also linked to the aetiology of diseases such as asthma, emphysema and heart disease. The clinical management of infection is problematic and antibiotic resistance is emerging. Increased understanding of immune processes that are involved in both clearance and immunopathology of chlamydial infection is critical for the development of improved treatment strategies. Here, we show that IL-13 was produced in the lungs of mice rapidly after Chlamydia muridarum (Cmu) infection and promoted susceptibility to infection. Wild-type (WT) mice had increased disease severity, bacterial load and associated inflammation compared to IL-13 deficient (−/−) mice as early as 3 days post infection (p.i.). Intratracheal instillation of IL-13 enhanced bacterial load in IL-13−/ − mice. There were no differences in early IFN-g and IL-10 expression between WT and IL-13−/ − mice and depletion of CD4+ T cells did not affect infection in IL-13−/ − mice. Collectively, these data demonstrate a lack of CD4+ T cell involvement and a novel role for IL-13 in innate responses to infection. We also showed that IL-13 deficiency increased macrophage uptake of Cmu in vitro and in vivo. Moreover, the depletion of IL-13 during infection of lung epithelial cells in vitro decreased the percentage of infected cells and reduced bacterial growth. Our results suggest that enhanced IL-13 responses in the airways, such as that found in asthmatics, may promote susceptibility to chlamydial lung infection. Importantly the role of IL-13 in regulating infection was not limited to the lung as we showed that IL-13 also promoted susceptibility to Cmu genital tract infection. Collectively our findings demonstrate that innate IL-13 release promotes infection that results in enhanced inflammation and have broad implications for the treatment of chlamydial infections and IL-13-associated diseases.
Introduction
Chlamydiae are Gram-negative, obligate intracellular bacteria that commonly cause respiratory and genital tract as well as ocular infections in humans. Globally, Chlamydophila pneumoniae has been estimated to account for 5% of cases of bronchitis and sinusitis, and up to 22% of cases of community-acquired pneumonia requiring hospitalisation [1], [2]. Chlamydia trachomatis is the world's most common sexually transmitted bacterial infection with an estimated 92 million new cases reported annually [3], and vertical transmission of C. trachomatis can initiate eye infections and pneumonia in new-borns [4], [5]. Chlamydiae commonly cause asymptomatic infections and significantly, between 50–80% and 10–20% of adults have anti-C. pneumoniae and anti-C. trachomatis antibodies respectively, indicating the high prevalence of these infections within the community [1], [6], [7], [8]. Furthermore, chlamydial infection has been linked with a number of chronic disease states including asthma [9], chronic obstructive pulmonary disease (COPD) [10], [11], atherosclerotic cardiovascular disease [12], and neurodegenerative disorders such as Alzheimer's disease [13]. Understanding the complex immunobiology of host-pathogen interactions and the delineation of the specific responses that drive clearance versus tissue damage are of paramount importance for the prevention and treatment of chlamydial infection and diseases.
CD4+ T helper type 1 (Th1) cells secreting IFN-γ play critical roles in the clearance of infection. The rate of clearance of Chlamydia from infected mouse lungs is directly proportional to increases in IFN-γ levels [14], [15], [16], and the absence of this cytokine or its receptor drives infection into a persistent state [17], [18], [19]. IFN-γ enhances the ability of macrophages to clear chlamydial infections [20], [21] and it is a powerful activator of indoleamine 2,3-dioxygenase (IDO) and inducible nitric oxide sythase (iNOS), which prevent bacterial growth by limiting tryptophan availability and upregulating nitric oxide (NO) production, respectively [22], [23], [24], [25]. Although Chlamydiae predominantly colonise epithelial cells, new evidence suggests that they can also infect smooth muscle cells, vascular endothelial cells and components of the immune system including macrophages and dendritic cells [26], [27]. Importantly, infection of alveolar macrophages of both human and mouse origin has been demonstrated and is associated with enhanced production of anti-inflammatory cytokines [28], [29], [30].
Severe forms of tissue damage due to Chlamydia infections of the respiratory and genital tracts are generally caused by infections that elicit both cytopathic and delayed type hypersensitivity immunopathologic destruction of the epithelium [31], [32]. In the case of C. pneumoniae and C. trachomatis this can lead to pneumonia and pelvic inflammatory disease and infertility, respectively. At the cell and molecular level immunopathology may consist of excessive infiltration of neutrophils, inflammatory monocytes, and the over-expression of the pro-inflammatory cytokines IL-1β and TNF-α [33], [34], [35].
Studies in animals have shown that a deficiency in the IFN-γ response, an overtly suppressive IL-10 response, or a delay in the development of a global T cell response during chlamydial infection can all lead to enhanced bacterial dissemination and disease sequelae [15], [16], [36]. Given these previous studies, it seems plausible that the development of anti-chlamydial Th2 immune responses could lead to increased disease susceptibility and immunopathology. In support of this concept, studies in our laboratory have shown that pulmonary infection with the natural mouse pathogen C. muridarum (Cmu) can lead to enhanced production of the Th2 cytokine IL-13. Cmu infection early in life leads to increased production of IL-13 following allergen challenge in adult mice [37], [38]. On a cellular level, Cmu infection of bone marrow-derived dendritic cells (BMDC) modulates the cytokine profile from both DCs and T cells to produce increased IL-13 in vitro [26]. Furthermore, urogenital Cmu infection in MyD88-deficient mice leads to a dominant Th2 response, skewed from the Th1/Th17 response of WT mice, which is associated with an ascending Cmu infection and severe pathology in the upper genital tract [39]. Allergic asthma is characterised by the infiltration of CD4+ Th type 2 (Th2) cells, which produce a specific subset of cytokines (e.g. IL-4, IL-5, IL-10 and IL-13) which have been linked to the pathogenesis of disease [40]. Clinical evidence directly links C. pneumoniae infection to asthma [41], [42], [43], [44]. However, the mechanistic basis underlying this relationship remains poorly understood. Enhanced production of IL-13 in response to chlamydial infection may contribute to the induction and exacerbation of asthma [26], [37], [38].
IL-13 has been described as a susceptibility factor for infection with both Leishmania major and Crytococcus neoformans [45], [46]. Both of these studies focus on the adaptive immune response to infection and attribute the ratio of Th2 cell derived IL-13 to Th1 cell IFN-γ as a key factor in determining the ability to mount a protective immune response and elimination of the pathogen. IL-13 is also produced by innate immune cells but the role of this cytokine in innate host defence against infection has received little attention. There is the potential that Chlamydia may induce innate and/or adaptive IL-13 responses to promote infection, which has significant implications for chlamydial respiratory and genital tract diseases, and associated conditions. In the present study we demonstrate that early production of IL-13 during the innate immune response plays a critical and previously unrecognised role in promoting Cmu infection of the respiratory and genital tract.
Results
Absence of IL-13 reduces susceptibility to and improves clearance of Cmu lung infection
We first assessed the role of IL-13 in chlamydial respiratory infection. Adult WT and IL-13−/ − mice were infected intranasally (i.n.) with Cmu and disease severity and bacterial numbers in the lungs were determined over time. Weight is an established indicator of disease severity, and substantial weight loss was observed in WT mice from 7 days after infection (Figure 1A). At this stage of infection mice begin to display significant histopathological changes in the lung [37]. By contrast, IL-13−/ − mice increased in body weight from day 7 p.i. (Figure 1A). These differences in weight change between WT and IL-13−/ − mice were significant from 9-20 days after infection (Figure 1A). Differences in weight changes between WT and IL-13−/ − mice also correlated with bacterial load in the lungs. WT mice had a significant increase in bacterial load from day 3 to day 15 p.i. (Figure 1B). By contrast, IL-13−/ − mice had significantly lower levels of Cmu at day 3 p.i. and throughout the course of infection (Figure 1B). The administration of recombinant mouse (rm)IL-13 to the lungs of IL-13−/ − mice prior to Cmu lung infection resulted in a significant increase in bacterial load at day 5 p.i. compared to untreated controls (Figure 1C). Together, these data demonstrate that the early presence of IL-13 during the innate host defence response plays a central role in promoting susceptibility to Cmu lung infection. Furthermore IL-13 deficiency suppresses the development of clinical signs infection as a result of an enhanced ability to clear the bacteria.
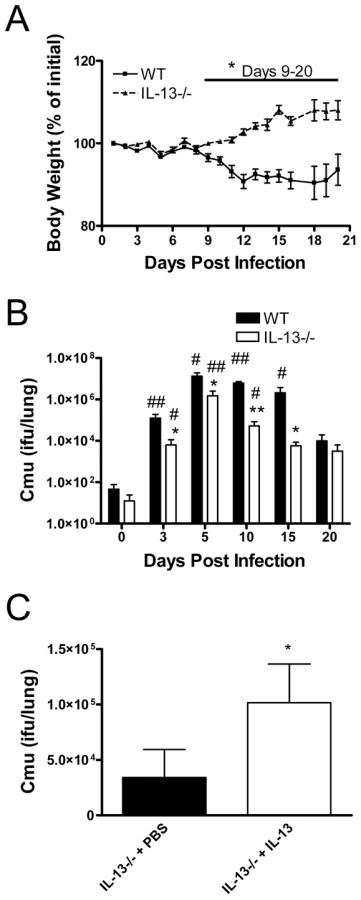
Enhanced Cmu clearance in the absence of IL-13 is associated with decreased airway inflammation
We then characterised the influence of IL-13 on inflammatory responses to Cmu infection. Pulmonary inflammation was assessed by enumerating leukocytes in the bronchoalveolar lavage fluid (BALF) of infected mice. Infection of WT mice led to a significant increase in the total number of leukocytes, neutrophils and macrophages present in the airways from as early as 5 days p.i. and significant increases in lymphocyte numbers were observed from 15 days p.i. (Figure 2A–D). IL-13 deficiency resulted in a significant reduction in total leukocyte, neutrophil, macrophage and lymphocyte influx into the lungs compared to WT controls, with limited evidence of increases in cellular infiltrates compared to baseline levels. (Figure 2A–D). Eosinophils were not detected in the airways of infected mice (not shown). Differences in neutrophil influx between strains were detected at 5 days p.i., whereas significant differences in macrophage and lymphocyte numbers were not apparent until the later stages of infection. An increased level of blood neutrophilia was also observed in infected WT compared to IL-13−/ − mice (data not shown). Therefore, IL-13 deficiency results in not only reduced bacterial load but decreases in associated inflammation.

IL-13 is expressed in the lungs during infection and influences the expression of cytokines important in pathogen clearance
Next we assessed the temporal expression of IL-13 during chlamydial infection. Quantitative real-time PCR of lung tissue RNA was used to measure the expression of IL-13 in WT mice following i.n. infection and normalised to uninfected controls. Infection was accompanied by a 5-fold increase in IL-13 expression, which was observed as early as 24 hours p.i. compared to naïve controls (Figure 3A). This increased level of IL-13 expression in the lungs of infected mice was maintained for at least 20 days p.i when compared to IL-13 expression in naïve tissue. These data represent the first report of pulmonary IL-13 expression during respiratory chlamydial infection.

To explore the mechanisms by which IL-13 mediates the host response to Cmu lung infection, we determined whether IL-13 influences the expression of IFN-γ and IL-10, factors known to play a central role in the immune response to chlamydial infection. Notably, the expression of IFN-γ and IL-10 were not different between naïve WT and IL-13−/ − mice and the onset of IFN-γ and IL-10 production was not affected by the absence of IL-13 during the early stages of infection. Indeed, on days 3 and 5 p.i. there were no significant differences between the expression of IFN-γ in infected IL-13−/ − mice compared to WT controls (Figure 3B). On day 10, IFN-γ production in WT mice was less than IL-13−/ − mice, however, by day 15 the production of IFN-γ was greater in WT mice. Interestingly, differences in the levels of IL-10 expression were not observed until day 15 p.i., where WT mice expressed more IL-10 compared to IL-13−/ − mice (Figure 3C). These results demonstrate that IL-13 directly or indirectly influences the expression of other cytokines that have been implicated in host defence against chlamydial infection. However, since these changes were not observed until the later stages of infection they do not explain the effect of IL-13 deficiency on bacterial numbers during the early onset of Cmu infection. This indicates that there are other mechanisms rather than changes in cytokine responses or in T cell phenotype that result in reduced infection in the absence of IL-13.
Enhanced clearance of Cmu respiratory infection in the absence of IL-13 is not dependent on CD4+ T-cells
The limited studies that have investigated the role of IL-13 in pathogen infection have focused on this molecule as a cytokine that is produced by activated CD4+ Th2 cells. By contrast, our data suggest that IL-13 plays an important role as early as 3 days p.i. (Figure 1B), and therefore is mediating innate rather than adaptive responses. To test this hypothesis we depleted CD4+ T cells using a specific monoclonal antibody (mAb) prior to and after i.n. infection of WT and IL-13−/ − mice. FACS analysis confirmed that antibody treatment depleted CD4+ cell numbers in the lung to less than 5% of that in untreated WT mice (0.75 ± 0.11 Vs. 13.91 ± 1.26% viable cells, Figure S1). WT mice treated with anti-CD4 mAb had increased chlamydial load in the lungs compared to untreated controls (Figure 4). Both untreated and treated WT mice displayed a significant drop in body weight 10 days post infection (93.17 ± 1.49 and 94.83 ± 1.78% of initial weight, p<0.01, respectively), which became apparent earlier in the treated group (data not shown). By contrast, CD4+ cell depletion did not significantly affect bacterial load (Figure 4) or body weight in IL-13−/ − mice (100.8 ± 0.70 and 100.57 ± 1.42% of initial weight in untreated and treated IL-13−/ − mice respectively). Together, these observations suggest that the reduced susceptibility to chlamydial infection in the absence of IL-13 is not mediated by CD4+ T cells but is linked to the innate host defence response.

Macrophages from IL-13 deficient mice show enhanced uptake of Cmu in vitro and in vivo
IL-13 is known to affect macrophage function and impair phagocytosis. Phagocytosis of bacteria by macrophages plays an important role in the innate defence against pathogens, including Chlamydia. Therefore, the effect of IL-13 deficiency on Cmu uptake by macrophages was investigated. Equal numbers of bone marrow-derived (BM) macrophages from WT and IL-13−/ − mice were cultured in the presence of equal titres of UV-inactivated, CFSE-labelled Cmu. Cells were then washed to remove any free Cmu and the percentage of BM macrophages (F4/80+ cells) that had taken up Cmu (CFSE+) was determined using flow cytometry. The percentage of Cmu positive macrophages was significantly higher in cultures from IL-13−/ − compared to WT mice (84.4 ± 2.3 Vs. 66.5 ± 1.1%, p<0.05, Figure 5A). Notably these methods directly measure phagocytosis rate of a specific number of Cmu by a specific number of macrophages and are not affected by the differing amounts of Cmu present in the two strains. To confirm these data in vivo, the effects of IL-13 on the function of macrophages during Cmu lung infection was also investigated. Alveolar macrophages were isolated from the BALF of infected WT and IL-13−/ − mice and the engulfment of Cmu was assessed by staining them using a Chlamydia-specific fluorescent labelling kit (Figure 5B). Interestingly, only half as many BALF macrophages from WT mice stained positive for Cmu 3 days after infection compared to those from IL-13−/ − mice (Figure 5C). This is despite total BALF macrophage numbers being similar (data not shown) and Cmu numbers increased in the lungs of WT compared to IL-13−/ − mice at this stage of infection (Figure 1B). At 5 days after infection no strain-specific differences in the ability of macrophages to engulf Cmu were detected, however, this may be explained by the high numbers of Cmu observed in the lungs of WT mice at this stage of infection. Together, our findings show that in the absence of IL-13, the uptake of Cmu by macrophages is enhanced. Thus, impaired phagocytosis of Cmu may represent an important mechanism by which chlamydial clearance is delayed in the presence of IL-13.

Depletion of IL-13 reduces the susceptibility of murine pulmonary epithelial cells to Cmu infection
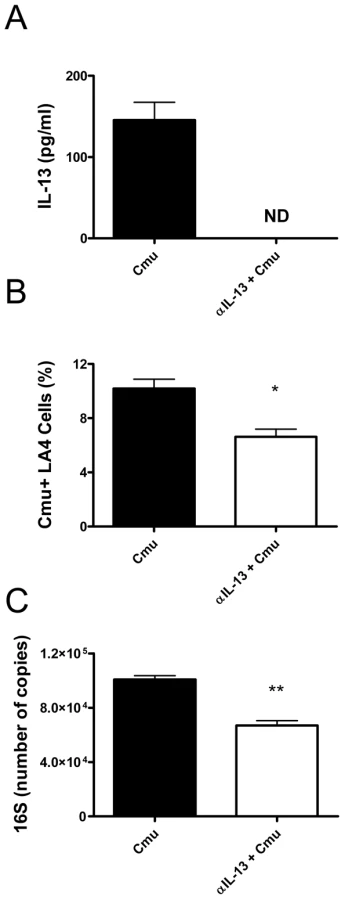
The absence of IL-13 had profound effects during the early stages of infection. Therefore, we hypothesised that, in addition to influencing macrophage phagocytosis, IL-13 may affect the susceptibility of airway epithelial cells to infection and intracellular proliferation of Cmu. To investigate this hypothesis we established an in vitro model of infection with LA4 cells, an immortalised murine lung epithelial cell line. Confluent monolayers of LA4 cells were infected with Cmu and cells were treated with anti-IL-13 (αIL-13) mAb prior to and during infection. Infection was assessed by enumerating Cmu inclusion positive cells using fluorescent microscopy and by determining the levels of Cmu 16S RNA expression in cultures. 16S expression is an indicator of growth. Previous work in our laboratory has shown that Cmu infection of LA4 cells induces widespread cell lysis after 30 hours (not shown). Therefore, the effects of IL-13 depletion on in vitro infection were assessed after 24 hours. Incubation for 24 hours still allows for the formation of large inclusions within infected cells and is appropriate for analysing of the effect of IL-13 depletion on cellular susceptibility to chlamydial infection. αIL-13 mAb treatment depleted IL-13 protein levels 24 hours after infection (Figure 6A). Significantly, the depletion of IL-13 resulted in decreased susceptibility of LA4 cells to infection. αIL-13 treated LA4 cultures had lower percentages of Cmu inclusion positive cells 24 hours after infection compared to untreated controls (Figure 6B). This observation was confirmed by quantitative real-time PCR analysis, demonstrating that IL-13 deficient cultures had lower copies of Cmu 16S than untreated cultures (Figure 6C). These results indicate that IL-13 directly promotes infection of lung epithelial cells by Cmu.
Absence of IL-13 reduces susceptibility to and improves clearance of chlamydial genital tract infection
To determine if IL-13 also played a role in promoting infection at other mucosal surfaces we intravaginally infected mice with Cmu and assessed disease severity and bacterial numbers over time. The chlamydial genital tract infection model employed in this study[47] has minimal effects on mouse body weight thus we used a clinical scoring system to determine disease severity (Table 1). Intravaginally infected WT mice had significantly higher clinical scores and more bacteria in vaginal lavage fluid 15 days p.i. compared to infected IL-13−/ − mice (Figure 7). These data suggest that the role of IL-13 in host responses to Chlamydia infection is not restricted to the lung and suggests that this cytokine may play a role in other Chlamydia associated diseases.


Discussion
In this study we have shown for the first time that IL-13 responses to Cmu infection are important in establishing and promoting chlamydial infection, and inflammation and disease in the respiratory and genital tracts. These effects are associated with reduced macrophage phagocytosis and enhanced infection of airway epithelial cells. Chlamydial respiratory and genital tract infections and diseases are prevalent throughout the world and infections are associated with a number of other diseases, particularly asthma. Elucidating the mechanisms that determine susceptibility to infection and chlamydial clearance may identify new ways of treating these conditions. We have previously demonstrated that Cmu infection in vitro and in vivo induces IL-13 responses [26], [38] and that transfer of Cmu infected murine bone marrow dendritic cells (BMDCs) into recipient mice subverts the in vivo immune response from a protective Th1 to a non-protective Th2 phenotype that may promote chronic infection [26]. In the present study we extend these studies and show that IL-13 promotes chlamydial infection and has an unexpected role in the immediate host defence responses to infection. Importantly, this association appears to be at the level of the innate rather than the adaptive immune response and is not predicated on alterations in T cells responses or the concomitant suppression of IFNγ responses.
In our study IL-13 was increased 5-fold in the lung as early as 24 hours after infection. IL-13 is a potent cytokine and low levels in the airway enhances infection (Figure 1C) and induces profound changes in lung physiology [48]. WT mice displayed increased symptoms, Cmu load and airway inflammatory cell burden after infection compared to their IL-13−/ − counterparts. Through their capacity to rapidly clear Cmu from the lungs IL-13−/ − mice circumvent the development of sequelae associated with chronic infection, including the influx of inflammatory cells into the airway. The role of IL-13 in infection is typically attributed to its function as a Th2 cytokine, often acting as an immunological ‘switch’ by downregulating the Th1 response. Indeed IL-13 has been identified as a susceptibility factor for infection of mice by the protozoan parasite Leishmania major by suppressing the expression of IFNγ and IL-12 [45]. Moreover, over-expression of IL-13 in transgenic mice enhanced pulmonary infection of mice with C. neoformans, which was associated with increased Th2 cytokine production [46]. Interestingly, there was no inverse relationship detected between Th1 and Th2 cytokine production in this setting; however, increased fungal load correlated with attenuated Th17 cytokine production 60 days p.i.
In our study the influence of IL-13 on chlamydial infection was evident as early as 3 days p.i., again suggesting a novel role for this cytokine in the innate rather than acquired immune response to infection. The level of Cmu in the lungs of CD4-depleted, IL-13−/ − mice was significantly lower than in both CD4-depleted, WT and untreated WT groups. Furthermore, there were no differences in IFN-g or IL-10 expression early in infection of WT and IL-13−/ − mice. These results demonstrate that protection against infection in IL-13−/ − mice is not dependent on CD4+ T-cell responses. These results confirm an important and central role for IL-13 in promoting infection during the early phases of the host defence response.
Chlamydia is capable of infecting a range of cell types, including alveolar macrophages and epithelial cells [27]. Our study demonstrates for the first time that the presence of IL-13 reduces the ability of macrophages to engulf Cmu in vitro and during in vivo lung infection. This may account for the enhanced ability of mice to clear this pathogen from the respiratory tract in the absence of this cytokine. Investigations of the role of IL-13 in immune responses to infection have shown that this molecule can induce the development of macrophages, which have a documented impairment in the ability to engulf and destroy intracellular pathogens in vitro [49], [50] and are associated with increased fungal burden in the lungs of mice during C. neoformans infection in vivo [46]. Interestingly, in vitro studies have also demonstrated that macrophage expression of the mannose receptor plays a pivotal role in determining susceptibility to chlamydial infection, although the significance of these findings in vivo have not yet been explored [51]. The capacity of IL-13 to modulate the innate immune response to infection in the present study may be underpinned by the development of macrophages with a reduced capacity to engulf and destroy Chlamydia in the lung.
It is also possible that IL-13 may condition the respiratory epithelium so that it is more susceptible to infection, thus the action of IL-13 is on structural cells as well as non-lymphoid cells that play key roles in host defence pathways. Furthermore, the fact that IL-13 mediates susceptibility to genital tract infection highlights the potential widespread role of this molecule in promoting chlamydial infection and diseases.
While CD4+ Th2 cells have typically been regarded as the principal source of IL-13, there is now a growing body of evidence that non-lymphoid cells are important producers of this cytokine and contribute to its associated pathologies. Mast cells [52], basophils [53], macrophages [54], bronchial mucosal cells [55], airway epithelial cells [56], dendritic cells [26] and natural killer T (NKT) cells [57] have all been demonstrated to generate IL-13. In our study IL-13 is produced rapidly, with an increase in expression apparent within 1 day p.i., which suggests that this cytokine is originating from non-lymphoid cells, with innate immune activity. Further studies are required to identify the key cell(s) that are the early cellular sources of IL-13 during Cmu infection.
The effects of IL-13 in promoting enhanced Cmu infection demonstrated in the current study may provide a basis for the widely observed clinical and experimental link between chlamydial infection and asthma [9], [38], [58], [59], [60]. Allergic airway inflammation in mice inhibits pulmonary host defence against other respiratory pathogens such as Pseudomonas aeruginosa [61] and alveolar macrophages from children with poorly controlled asthma have an impaired ability to phagocytose FITC-conjugated Staphylococcus aureus [62]. Furthermore BALB/c mice, which are biased towards Th2 cytokine responses, are markedly more susceptible to chlamydial lung infection than the Th1-predisposed C57BL/6 strain [16]. Increases in pulmonary IL-13 in asthmatic patients may promote susceptibility and contribute to the prevalence of chlamydial infection in these patient populations. Our in vitro evidence that IL-13 increases the susceptibility of airway epithelial cells to infection with Cmu also supports this concept.
In summary, our study reveals for the first time that production of IL-13 during the innate host defence phase plays a central role in establishing and promoting Cmu respiratory and genital tract infections. This role appears to be independent of CD4+ T cell-mediated adaptive immune responses and may be a result of the reduced ability of macrophages to engulf Cmu and an increased susceptibility of pulmonary epithelial cells to infection. This study enhances our understanding of the pathogenesis of chlamydial infection and identifies IL-13 as new potential target to attenuate infection, inflammation and pathology associated with Chlamydia.
Materials and Methods
Ethics statement
This study was carried out in strict accordance with the recommendations set out in the Australian code of practice for the care and use of animals for scientific purposes issued by the National Health and Medical Research Council (Australia). All protocols were approved by the University of Newcastle Animal Care and Ethics Committee and all efforts were made to minimise suffering.
Mice
Adult (6–8 weeks old) WT BALB/c mice and IL-13−/ − mice on a BALB/c background were supplied by the animal breeding facilities of the Australian National University or the University of Newcastle. Mice were housed under specific pathogen free conditions.
Respiratory tract infection
Adult mice were infected i.n. with 100 ifu of Cmu (ATCC VR-123; in 30 µl of sucrose phosphate glutamate buffer [SPG])[37], [38], [60], [63]. Mice were monitored over a 20 day period and rate of weight gain/loss was used as a measure of clinical condition. At selected time points, mice were sacrificed by sodium pentobarbital overdose (Abbott Australasia, Kurnell, Australia) for analysis.
Recombinant IL-13 administration
IL-13−/ − mice were intratracheally administered rmIL-13 (10 ng, 30 µl of PBS, R&D Systems, Gymea, NSW) or PBS 6 hours prior to infection and sacrificed 5 days later for analysis.
Genital tract infection
Adult mice were subcutaneously injected with 2 mg medroxyprogesterone acetate (Troy Laboratories, Smithfield, Australia) to synchronise their estrous cycles. Seven days later mice were infected intravaginally with 5×104 ifu Cmu (in 20 µl SPG)[47]. Mice were monitored over a 15 day period and their clinical score determined according to specific signs of disease (Table 1). At selected time points, mice were sacrificed by sodium pentobarbital overdose for analysis.
Assessment of bacterial levels
Whole lungs from mice infected i.n. with Cmu were removed and stored at −80°C. Vaginal lavage was performed on intravaginally infected mice by flushing the vaginal vault with 2×60 µl Hanks buffered salt solution (HBSS; Trace Scientific, Noble Park, NSW). DNA extractions were performed and Cmu numbers (IFU) determined in whole lungs or vaginal lavage by real-time quantitative PCR and comparison with known standards as previously described [37], [38], [60], [63].
Airway inflammation
BALF was obtained by cannulation of the trachea and flushing the airways with 2× 1 ml HBSS [37]. BALF cytospins were stained with May-Grunwald-Giemsa and leukocytes enumerated by morphological criteria (≈300 cells by light microscopy [40X]) [37]. All samples were coded and counts performed in a blinded fashion.
Gene expression analysis
Cytokine expression was evaluated by real-time PCR [38], [60]. Total RNA was extracted from all samples using TRIZOL according to the manufacturer's instructions (Invitrogen, Mount Waverley, VIC). Reverse transcription of RNA (1000 ng) was performed using Superscript III and random hexamer primers (Invitrogen). Relative abundance of genes was determined compared to the reference gene hypoxanthine-guanine phosphoribosyltransferase using a Prism7000 Sequence Detection System (Applied Biosystems, Scoresby, VIC). Primers used were; IFN-γ, Fwd 5′ - TCT TGA AAG ACA ATC AGG CCA TCA, Rev 3′-, GAA TCA GCA GCG ACT CCT TTT CC, IL-10, Fwd 5′ - CAT TTG AAT TCC CTG GGT GAG AAG, Rev 3′-, GCC TTG TAG ACA CCT TGG TCT TGG, IL-13 Fwd 5′ - AGC TGA GCA ACA TCA CAC AAG ACC, Rev 3′-, TGG GCT ACT TCG ATT TTG GTA TCG, 16S of Cmu, Fwd 5′ - GCG GCA GAA ATG TCG TTT T, Rev 3′-, CGC TCG TTG CGG GAC TTA and hypoxanthine-guanine phosphoribosyltransferase, Fwd 5′ - AGG CCA GAC TTT GTT GGA TTT GAA, Rev 5′ - CAA CTT GCG CTC ATC TTA GGC TTT.
CD4+ T-cell depletion
Mice were treated intraperitoneally with 300 µg αCD4 (clone GK1.5) on days -3, -1, 2 and 5 of Cmu lung infection. The effect of T-cell depletion on bacterial recovery was assessed in whole lungs on day 10. Depletion of CD4+ T-cells was confirmed on day 10 by flow cytometry (Figure S1).
Assessment of in vitro uptake of Cmu by BM macrophages
Femurs and tibias of WT and IL-13−/ − mice were collected and bone marrow flushed out with complete RPMI (RPMI 1640, 5×10−5 M 2-mercaptoethanol, 10% heat-inactivated FCS, 2 mM L-glutamine, 20 mM HEPES, 100 µg/ml penicillin and 100 µg/ml streptomycin). Red blood cells were lysed and cells washed through a 70 µm nylon cell strainer. The cells were plated out at 1×105 cells/ml in 10 mls complete RPMI supplemented with 15 ng/ml rmGM-CSF (Gift from Walter and Eliza Hall Institute [WEHI], Melbourne) and incubated at 37°C 5% CO2. On day 3, another 5 ml medium containing 15 ng/ml rmGM-CSF was added. On day 7, supernatants were removed and non-adherent and semi-adherent cells were removed by washing in cold PBS. Adherent cells were recovered by gentle cell scraping and washed using cold PBS supplemented with 2 mM EDTA. These cells were shown to be >90% macrophages by flow cytometry. BM macrophages were plated out (2×105 cells/ml, 96 well plate) and inoculated with CFSE-labelled, UV-inactivated Cmu (MOI 5) and incubated overnight. (37°C, 5% CO2) in complete RPMI. Cultures were washed with complete RPMI to remove all free Cmu and the percentage of BM macrophages (F4/80+ cells) that stained positive for Cmu (CFSE+F4/80+ cells) was determined by flow cytometry.
Assessment of in vivo uptake of Cmu by lung macrophages
BALF was collected from mice on days 3 and 5 following respiratory tract infection. BALF cells were incubated on sterile 10 mm round glass coverslips in a 48 well culture plate for 1 h (37°C, 5% CO2) in complete RPMI to allow adhesion of macrophages. Coverslips were removed and adhered cells stained using the Chlamydia Cel LPS kit (CelLabs, Brookvale, NSW) according to the manufacturers instructions. The percentage of macrophages that stained positive for the presence of Chlamydia was determined in each sample (≈300 macrophages assessed by fluorescent microscopy [40X]). All samples were coded and counts performed in a blinded fashion.
In vitro Cmu infection of mouse lung epithelial cells
LA4 cells (3×105) were plated out on sterile 10 mm glass coverslips in a 48 well culture plate and incubated in Iscove's modified Eagle media (500 µl, 10% fetal calf serum) for 24 h in the presence of either 100 µg αIL-13 mAb (R&D Systems) or PBS (untreated control). Confluent cell monolayers were then infected with Cmu (MOI 5), treated again with αIL-13 mAb or PBS and cultured for a further 24 h. Supernatants were collected and IL-13 ELISA performed according to the manufacturer's instructions (R&D Systems). For staining of chlamydial inclusions, coverslips were removed and cells stained with the Chlamydia Cel LPS kit. The percentage of Cmu inclusion positive cells was determined for each treatment (average of >10 fields determined at 40× magnification using a fluorescent microscope). To determine 16S Cmu RNA expression RNA was prepared and assayed by real-time PCR as described above.
Statistical analysis
Results are presented as mean±SEM. Statistical significance of whole data sets was initially confirmed using one-way ANOVA. The Wilcoxon Rank-sum test was used for non-parametric tests (Mann-Whitney test for two independent samples). P<0.05 was considered statistically significant.
Supporting Information
{kind=link}
Zdroje
1. KuoCC
JacksonLA
CampbellLA
GraystonJT
1995 Chlamydia pneumoniae (TWAR). Clin Microbiol Rev 8 451 461
2. BlasiF
2004 Atypical pathogens and respiratory tract infections. Eur Respir J 24 171 182
3. World Health Organisation, Department of HIV/AIDS 2001 Global prevalence and incidence of selected curable sexually transmitted infections: Overview and estimates. Available: http://whqlibdoc.who.int/hq/2001/WHO_HIV_AIDS_2001.02.pdf Accessed 17 March 2011
4. JainS
1999 Perinatally acquired Chlamydia trachomatis associated morbidity in young infants. J Matern Fetal Med 8 130 133
5. RoursGI
HammerschlagMR
Van DoornumGJ
HopWC
de GrootR
2009 Chlamydia trachomatis respiratory infection in Dutch infants. Arch Dis Child 94 705 707
6. GraystonJT
1992 Infections caused by Chlamydia pneumoniae strain TWAR. Clin Infect Dis 15 757 761
7. LyytikainenE
KaasilaM
KoskelaP
LehtinenM
PatamaT
2008 Chlamydia trachomatis seroprevalence atlas of Finland 1983-2003. Sex Transm Infect 84 19 22
8. SmithJS
MunozN
HerreroR
Eluf-NetoJ
NgelangelC
2002 Evidence for Chlamydia trachomatis as a human papillomavirus cofactor in the etiology of invasive cervical cancer in Brazil and the Philippines. J Infect Dis 185 324 331
9. HansbroPM
BeagleyKW
HorvatJC
GibsonPG
2004 Role of atypical bacterial infection of the lung in predisposition/protection of asthma. Pharmacol Ther 101 193 210
10. BrandenE
GnarpeJ
HillerdalG
OrreL
SkoldCM
2007 Detection of Chlamydia pneumoniae on cytospin preparations from bronchoalveolar lavage in COPD patients and in lung tissue from advanced emphysema. Int J Chron Obstruct Pulmon Dis 2 643 650
11. BrandenE
KoyiH
GnarpeJ
GnarpeH
TornlingG
2005 Chronic Chlamydia pneumoniae infection is a risk factor for the development of COPD. Respir Med 99 20 26
12. SessaR
NicolettiM
Di PietroM
SchiavoniG
SantinoI
2009 Chlamydia pneumoniae and atherosclerosis: current state and future prospectives. Int J Immunopathol Pharmacol 22 9 14
13. HonjoK
van ReekumR
VerhoeffNPLG
2009 Alzheimer's disease and infection: Do infectious agents contribute to progression of Alzheimer's disease? Alzheimers Dement 5 348 360
14. WangS
FanY
BrunhamRC
YangX
1999 IFN-γ knockout mice show Th2-associated delayed-type hypersensitivity and the inflammatory cells fail to localize and control chlamydial infection. Eur J Immunol 29 3782 3792
15. YangX
GartnerJ
ZhuL
WangS
BrunhamRC
1999 IL-10 gene knockout mice show enhanced Th1-like protective immunity and absent granuloma formation following Chlamydia trachomatis lung infection. J Immunol 162 1010 1017
16. YangX
HayGlassKT
BrunhamRC
1996 Genetically determined differences in IL-10 and IFN-gamma responses correlate with clearance of Chlamydia trachomatis mouse pneumonitis infection. J Immunol 156 4338 4344
17. ItoJI
LyonsJM
1999 Role of gamma interferon in controlling murine chlamydial genital tract infection. Infect Immun 67 5518 5521
18. RottenbergME
Gigliotti RothfuchsAC
GigliottiD
SvanholmC
BandholtzL
1999 Role of innate and adaptive immunity in the outcome of primary infection with Chlamydia pneumoniae, as analyzed in genetically modified mice. J Immunol 162 2829 2836
19. BeattyWL
ByrneGI
MorrisonRP
1993 Morphologic and antigenic characterization of interferon gamma-mediated persistent Chlamydia trachomatis infection in vitro. Proc Nat Acad Sci 90 3998 4002
20. ZhongGM
de la MazaLM
1988 Activation of mouse peritoneal macrophages in vitro or in vivo by recombinant murine gamma interferon inhibits the growth of Chlamydia trachomatis serovar L1. Infect Immun 56 3322 3325
21. ChenB
StoutR
CampbellWF
1996 Nitric oxide production: a mechanism of Chlamydia trachomatis inhibition in interferon-gamma-treated RAW264.7 cells. FEMS Immunol Med Microbiol 14 109 120
22. HassanainHH
ChonSY
GuptaSL
1993 Differential regulation of human indoleamine 2,3-dioxygenase gene expression by interferons-gamma and -alpha. Analysis of the regulatory region of the gene and identification of an interferon-gamma-inducible DNA-binding factor. J Biol Chem 268 5077 5084
23. MacMickingJ
XieQW
NathanC
1997 Nitric oxide and macrophage function. Annu Rev Immunol 15 323 350
24. MehtaSJ
MillerRD
RamirezJA
SummersgillJT
1998 Inhibition of Chlamydia pneumoniae replication in HEp-2 cells by interferon-gamma: role of tryptophan catabolism. J Infect Dis 177 1326 1331
25. IgietsemeJU
PerryLL
AnanabaGA
UririIM
OjiorOO
1998 Chlamydial infection in inducible nitric oxide synthase knockout mice. Infect Immun 66 1282 1286
26. KaikoGE
PhippsS
HickeyDK
LamCE
HansbroPM
2008 Chlamydia muridarum infection subverts dendritic cell function to promote Th2 immunity and airways hyperreactivity. J Immunol 180 2225 2232
27. BeagleyKW
HustonWM
HansbroPM
TimmsP
2009 Chlamydial infection of immune cells: altered function and implications for disease. Crit Rev Immunol 29 275 305
28. RedeckeV
DalhoffK
BohnetS
BraunJ
MaassM
1998 Interaction of Chlamydia pneumoniae and human alveolar macrophages: infection and inflammatory response. Am J Respir Cell Mol Biol 19 721 727
29. GaydosCA
SummersgillJT
SahneyNN
RamirezJA
QuinnTC
1996 Replication of Chlamydia pneumoniae in vitro in human macrophages, endothelial cells, and aortic artery smooth muscle cells. Infect Immun 64 1614 1620
30. HaranagaS
YamaguchiH
IkejimaH
FreidmanH
YY
2003 Chlamydia pneumoniae infection of alveolar macrophages: A model. J Infect Dis 187 1107 1115
31. DebattistaJ
TimmsP
AllanJ
AllanJ
2003 Immunopathogenesis of Chlamydia trachomatis infections in women. Fertil Steril 79 1273 1287
32. DarvilleT
HiltkeTJ
2010 Pathogenesis of genital tract disease due to Chlamydia trachomatis. J Infect Dis 201 Suppl 2 S114 125
33. LeeHY
SchripsemaJH
SigarIM
MurrayCM
LacySR
2010 A link between neutrophils and chronic disease manifestations of Chlamydia muridarum urogenital infection of mice. FEMS Immunol Med Microbiol 59 108 116
34. HvidM
BaczynskaA
DeleuranB
FedderJ
KnudsenHJ
2007 Interleukin-1 is the initiator of fallopian tube destruction during Chlamydia trachomatis infection. Cell Microbiol 9 2795 2803
35. SkworTA
AtikB
KandelRP
AdhikariHK
SharmaB
2008 Role of secreted conjunctival mucosal cytokine and chemokine proteins in different stages of trachomatous disease. PLoS Negl Trop Dis 2 e264
36. WangC
van GinkelFW
KimT
LiD
LiY
2008 Temporal delay of peak T-cell immunity determines Chlamydia pneumoniae pulmonary disease in mice. Infect Immun 76 4913 4923
37. HorvatJC
BeagleyKW
WadeMA
PrestonJA
HansbroNG
2007 Neonatal chlamydial infection induces mixed T-cell responses that drive allergic airway disease. Am J Respir Crit Care Med 176 556 564
38. HorvatJC
StarkeyMR
KimRY
PhippsS
GibsonPG
2010 Early-life chlamydial lung infection enhances allergic airways disease through age-dependent differences in immunopathology. J Allergy Clin Immunol 125 617 625, 625 e611-625 e616
39. ChenL
LeiL
ChangX
LiZ
LuC
2010 Mice deficient in MyD88 Develop a Th2-dominant response and severe pathology in the upper genital tract following Chlamydia muridarum infection. J Immunol 184 2602 2610
40. HansbroPM
KaikoGE
FosterPS
2011 Cytokine/anti-cytokine therapy - novel treatments for asthma? Br J Pharmacol In Press
41. CunninghamAF
JohnstonSL
JuliousSA
LampeFC
WardME
1998 Chronic Chlamydia pneumoniae infection and asthma exacerbations in children. Eur Respir J 11 345 349
42. BlackPN
ScicchitanoR
JenkinsCR
BlasiF
AllegraL
2000 Serological evidence of infection with Chlamydia pneumoniae is related to the severity of asthma. Eur Respir J 15 254 259
43. WarkPAB
JohnstonSL
SimpsonJL
HensleyMJ
GibsonPG
2002 Chlamydia pneumoniae immunoglobulin A reactivation and airway inflammation in acute asthma. Eur Respir J 20 834 840
44. WebleyWC
TilahunY
LayK
PatelK
StuartES
2009 Occurrence of Chlamydia trachomatis and Chlamydia pneumoniae in paediatric respiratory infections. Eur Respir J 33 360 367
45. MatthewsDJ
EmsonCL
McKenzieGJ
JolinHE
BlackwellJM
2000 IL-13 is a susceptibility factor for Leishmania major infection. J Immunol 164 1458 1462
46. MullerU
StenzelW
KohlerG
WernerC
PolteT
2007 IL-13 induces disease-promoting type 2 cytokines, alternatively activated macrophages and allergic inflammation during pulmonary infection of mice with Cryptococcus neoformans. J Immunol 179 5367 5377
47. BerryLJ
HickeyDK
SkeldingKA
BaoS
RendinaAM
2004 Transcutaneous immunization with combined cholera toxin and CpG adjuvant protects against Chlamydia muridarum genital tract infection. Infect Immun 72 1019 1028
48. YangM
HoganSP
HenryPJ
MatthaeiKI
McKenzieAN
2001 Interleukin-13 mediates airways hyperreactivity through the IL-4 receptor-alpha chain and STAT-6 independently of IL-5 and eotaxin. Am J Respir Cell Mol Biol 25 522 530
49. VarinA
MukhopadhyayS
HerbeinG
GordonS
2010 Alternative activation of macrophages by IL-4 impairs phagocytosis of pathogens but potentiates microbial-induced signalling and cytokine secretion. Blood 115 353 362
50. HarrisJ
De HaroSA
MasterSS
KeaneJ
RobertsEA
2007 T helper 2 cytokines inhibit autophagic control of intracellular Mycobacterium tuberculosis. Immunity 27 505 517
51. KuoCC
PuolakkainenM
LinTM
WitteM
CampbellLA
2002 Mannose-receptor positive and negative mouse macrophages differ in their susceptibility to infection by Chlamydia species. Microb Pathog 32 43 48
52. GessnerA
MohrsK
MohrsM
2005 Mast cells, basophils, and eosinophils acquire constitutive IL-4 and IL-13 transcripts during lineage differentiation that are sufficient for rapid cytokine production. J Immunol 174 1063 1072
53. SchroederJT
2009 Basophils beyond effector cells of allergic inflammation. Adv Immunol 101 123 161
54. KimEY
BattaileJT
PatelAC
YouY
AgapovE
2008 Persistent activation of an innate immune response translates respiratory viral infection into chronic lung disease. Nat Med 14 633 640
55. KotsimbosTC
ErnstP
HamidQA
1996 Interleukin-13 and interleukin-4 are coexpressed in atopic asthma. Proc Assoc Am Physicians 108 368 373
56. TemannUA
RayP
FlavellRA
2002 Pulmonary overexpression of IL-9 induces Th2 cytokine expression, leading to immune pathology. J Clin Invest 109 29 39
57. AkbariO
StockP
MeyerE
KronenbergM
SidobreS
2003 Essential role of NKT cells producing IL-4 and IL-13 in the development of allergen-induced airway hyperreactivity. Nat Med 9 582 588
58. MetzG
KraftM
2010 Effects of atypical infections with Mycoplasma and Chlamydia on asthma. Immunol Allergy Clin North Am 30 575 585, vii-viii
59. WebleyWC
SalvaPS
AndrzejewskiC
CirinoF
WestCA
2005 The bronchial lavage of pediatric patients with asthma contains infectious Chlamydia. Am J Respir Crit Care Med 171 1083 1088
60. HorvatJC
StarkeyMR
KimRY
BeagleyKW
PrestonJA
2010 Chlamydial respiratory infection during allergen sensitization drives neutrophilic allergic airways disease. J Immunol 184 4159 4169
61. BeisswengerC
KandlerK
HessC
GarnH
FelgentreffK
2006 Allergic airway inflammation inhibits pulmonary antibacterial host defense. J Immunol 177 1833 1837
62. FitzpatrickAM
HolguinF
TeagueWG
BrownLA
2008 Alveolar macrophage phagocytosis is impaired in children with poorly controlled asthma. J Allergy Clin Immunol 121 1372 1378, 1378 e1371-1373
63. SkeldingKA
HickeyDK
HorvatJC
BaoS
RobertsKG
2006 Comparison of intranasal and transcutaneous immunization for induction of protective immunity against Chlamydia muridarum respiratory tract infection. Vaccine 24 355 366
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2011 Číslo 5
- Farmakovigilanční studie perorálních antivirotik indikovaných v léčbě COVID-19
- Jak souvisí postcovidový syndrom s poškozením mozku?
- Měli bychom postcovidový syndrom léčit antidepresivy?
- 10 bodů k očkování proti COVID-19: stanovisko České společnosti alergologie a klinické imunologie ČLS JEP
Nejčtenější v tomto čísle
- Crystal Structure and Functional Analysis of the SARS-Coronavirus RNA Cap 2′-O-Methyltransferase nsp10/nsp16 Complex
- Lymphoadenopathy during Lyme Borreliosis Is Caused by Spirochete Migration-Induced Specific B Cell Activation
- The OXI1 Kinase Pathway Mediates -Induced Growth Promotion in Arabidopsis
- : An Emerging Cause of Sexually Transmitted Disease in Women