
Transcription Is Required to Establish Maternal Imprinting at the Prader-Willi Syndrome and Angelman Syndrome Locus
The Prader-Willi syndrome (PWS [MIM 17620]) and Angelman syndrome (AS [MIM 105830]) locus is controlled by a bipartite imprinting center (IC) consisting of the PWS-IC and the AS-IC. The most widely accepted model of IC function proposes that the PWS-IC activates gene expression from the paternal allele, while the AS-IC acts to epigenetically inactivate the PWS-IC on the maternal allele, thus silencing the paternally expressed genes. Gene order and imprinting patterns at the PWS/AS locus are well conserved from human to mouse; however, a murine AS-IC has yet to be identified. We investigated a potential regulatory role for transcription from the Snrpn alternative upstream exons in silencing the maternal allele using a murine transgene containing Snrpn and three upstream exons. This transgene displayed appropriate imprinted expression and epigenetic marks, demonstrating the presence of a functional AS-IC. Transcription of the upstream exons from the endogenous locus correlates with imprint establishment in oocytes, and this upstream exon expression pattern was conserved on the transgene. A transgene bearing targeted deletions of each of the three upstream exons exhibited loss of imprinting upon maternal transmission. These results support a model in which transcription from the Snrpn upstream exons directs the maternal imprint at the PWS-IC.
Published in the journal:
. PLoS Genet 7(12): e32767. doi:10.1371/journal.pgen.1002422
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002422
Summary
The Prader-Willi syndrome (PWS [MIM 17620]) and Angelman syndrome (AS [MIM 105830]) locus is controlled by a bipartite imprinting center (IC) consisting of the PWS-IC and the AS-IC. The most widely accepted model of IC function proposes that the PWS-IC activates gene expression from the paternal allele, while the AS-IC acts to epigenetically inactivate the PWS-IC on the maternal allele, thus silencing the paternally expressed genes. Gene order and imprinting patterns at the PWS/AS locus are well conserved from human to mouse; however, a murine AS-IC has yet to be identified. We investigated a potential regulatory role for transcription from the Snrpn alternative upstream exons in silencing the maternal allele using a murine transgene containing Snrpn and three upstream exons. This transgene displayed appropriate imprinted expression and epigenetic marks, demonstrating the presence of a functional AS-IC. Transcription of the upstream exons from the endogenous locus correlates with imprint establishment in oocytes, and this upstream exon expression pattern was conserved on the transgene. A transgene bearing targeted deletions of each of the three upstream exons exhibited loss of imprinting upon maternal transmission. These results support a model in which transcription from the Snrpn upstream exons directs the maternal imprint at the PWS-IC.
Introduction
Genomic imprinting is an epigenetic phenomenon that occurs at a subset of chromosomal regions and results in parent-of-origin specific monoallelic gene expression. Imprinted genes are frequently found in clusters, coordinately regulated by imprinting centers (ICs) that direct allele-specific differences in transcription, DNA methylation, histone modifications and replication timing [1]–[4]. Appropriate control of imprinted gene expression is vital to growth and development and errors in imprinting may lead to developmental disorders or embryonic lethality.
Prader-Willi syndrome (PWS) and Angelman syndrome (AS) are distinct neurogenetic disorders resulting from improper gene expression from an imprinted domain on chromosome 15q11–q13, the PWS/AS locus. Mutations arising from the maternal chromosome that lead to a loss of UBE3A function are sufficient to cause AS [5], [6]. PWS results from the loss of multiple paternal gene products encoded at the PWS/AS locus. A bipartite IC, comprised of the PWS-IC and the AS-IC, regulates both epigenetic reprogramming and allele-specific gene expression at the PWS/AS locus [7], [8]. Prevailing models of IC function suggest that the PWS-IC is a positive element that activates gene expression from the paternal allele. The AS-IC acts as a negative element to direct inhibitory epigenetic modifications at the PWS-IC during oogenesis, thereby silencing the paternally expressed genes on the future maternal allele [8]–[10]. A subset of individuals with PWS or AS bear microdeletions that disrupt gene expression at the PWS/AS locus. The shortest regions of overlap for these microdeletions define the boundaries of the IC. The PWS-IC is located in a region of 4.3 kb, just 5′ to and including exon 1 of SNRPN, while the AS-IC is contained within 0.88 kb approximately 35 kb upstream of the PWS-IC [11], [12]. Notably, within the 0.88 kb of AS-IC sequence reside two of several alternative upstream exons of SNRPN [13], [14]. These upstream exons are postulated to play a role in silencing the PWS-IC on the maternal allele [8], [15].
The PWS/AS locus is highly conserved from human to mouse in both gene order and allelic gene expression patterns, providing an excellent model for studying IC-directed imprinting mechanisms within this domain (Figure 1A). Hindering these studies has been the absence of an identifiable murine AS-IC. However, as in the human, the murine Snrpn locus contains several alternative upstream promoters and exons, herein referred to as U exons, from which multiple alternatively spliced transcripts arise [16], [17]. These U exons are transcribed exclusively from the paternal allele in the brain as well as in oocytes [15], [16], [18]. U exon expression in the oocyte suggests that transcription is essential for maternal imprinting at the PWS/AS locus. In this report, we test whether the murine U exons have AS-IC activity.

Previous targeted deletion approaches to knockout AS-IC function proved unsuccessful or resulted in only partial imprinting defects [19], [20], suggesting that the murine AS-IC consists of multiple elements. We have therefore taken a transgenic approach to identify the murine AS-IC. We utilized a BAC transgene that contains Snrpn and 100 kb of upstream sequence, including three of the alternative U exons. We show that this transgene is properly imprinted in multiple transgenic lines, indicating that there is a functional AS-IC contained within its sequence. Transcription of the U exons from the transgene correlates with imprint establishment in the maternal germ line. Upon targeted deletion of the three U exons, this transgene loses both the epigenetic imprint as well as its imprinted expression pattern, demonstrating that the AS-IC activity originates from the U exons.
Results
AS-IC Activity Is Contained within a 100-kb Region Upstream of Snrpn
To establish a transgenic system that would faithfully model AS-IC activity, it was necessary to identify a murine BAC that contained the PWS-IC and expressed Snrpn exclusively after paternal transmission. In addition, to study the role of the U exons, it was important to compare BAC transgenes containing U exons to those without. We screened the C57BL/6 RPCI-23 murine BAC library and identified BACs 380J10 and 425D18 for further study. The 380J10 BAC contains approximately 15 kb of sequence upstream of Snrpn exon 1, a region lacking any alternative Snrpn U exons. The 425D18 BAC possesses approximately 100 kb of sequence upstream of Snrpn, including three of the nine identified U exons (Figure 1B). We generated two independent transgenic lines for the 380J10 BAC, 380A and 380D. 380A contains multiple copies of the transgene while 380D is a single copy line (Figure S1A). We examined the imprinted status of the transgene by Northern blot analysis on postnatal (P) day 1 brain RNA. As the endogenous Snrpn transcript would complicate transgene expression analysis, we initially crossed our transgenic lines to a mutant bearing a 35 kb deletion of the PWS-IC (PWS-ICΔ35kb) [21]. This deletion ablates endogenous Snrpn expression when inherited paternally. The 380A and 380D lines were analyzed upon both maternal and paternal transmission of the transgene in combination with a paternal PWS-ICΔ35kb allele. In both lines, transgene-encoded Snrpn was expressed after maternal and paternal transmission (Figure 2A). This result demonstrated that AS-IC activity is not contained within 15 kb upstream of Snrpn, a region deficient in U exons.

We then focused on the 425D18 BAC which contains three Snrpn U exons. To simplify transgene expression analysis, we utilized BAC recombineering techniques to delete a region between Snrpn exons 5 and 7, sequence shown previously to have no effect on imprinting at the locus [21]. We established multiple transgenic lines using this BAC, termed BAC 425Δ5-7. RT-PCR with primers located in exons 4 and 8, the exons flanking the deletion, distinguishes expression of endogenous Snrpn from expression of the transgene based on amplicon size (Figure 3A). Analysis of P1 brain RNA showed the 425Δ5-7 transgene was expressed upon paternal but not maternal inheritance and thus was correctly imprinted (Figure 3B). This result was demonstrated in three independent lines, lines A, H, and I, indicating that AS-IC activity lies within the 425D18 BAC sequence. We chose line 425Δ5-7A for further analysis as we found this line to be single copy (Figure S1B).

We further analyzed imprinting of the 425Δ5-7 transgene by examining the Snrpn differentially methylated region (DMR), which is located within the PWS-IC. In somatic cells, this DMR is methylated on the maternal chromosome and unmethylated on the paternal chromosome [22]–[24]. This DNA methylation imprint is erased in fetal germ cells and applied as a maternal-specific epigenetic mark during oogenesis [25]–[27]. To investigate whether the 425Δ5-7 BAC transgene undergoes appropriate epigenetic reprogramming in the germ line, we used genomic bisulfite sequence analysis to examine the methylation status of the Snrpn DMR in transgenic P1 brain DNA. We bred the 425Δ5-7A transgene onto a C57BL/6 line congenic for the PWS/AS domain of Mus musculus castaneus chromosome 7 (B6.cast.c7) [28] and examined a region within the DMR containing 14 CpG dinucleotides. A single nucleotide polymorphism (SNP) located within our region of interest distinguishes C57BL/6 transgenic from endogenous cast sequences. As expected, the paternally transmitted 425Δ5-7A transgene displayed hypomethylation at the Snrpn DMR while the maternally transmitted transgene was hypermethylated, demonstrating the presence of the appropriate epigenetic imprinting marks (Figure 3C). The endogenous alleles displayed the expected mixture of methylated and unmethylated clones as both the maternal and paternal alleles are represented in these sequences (Figure S2). Taken together, the expression analysis and DNA methylation analysis illustrate that the 425D18 BAC harbors complete AS-IC activity.
Snrpn U Exon Expression Patterns Are Conserved on the 425Δ5-7 Transgene
The human AS-IC lies within a region of 880 bp that contains two of several alternative upstream exons of SNRPN, designated u5 and u6 [11], [13], [14], [29], [30]. These two exons are included in various transcripts originating from the upstream exons, suggesting a potential role in the imprinting process. A conserved sequence from the human AS-IC has not been identified in the mouse but the murine Snprn locus does possess a number of alternative U exons. Nine have been identified to date, termed U1-U9, spanning over 580 kb upstream of Snrpn exon 1 [17]. These exons generate multiple transcripts, most frequently splicing into exon 2 of Snrpn but occasionally splicing downstream of the Snrpn gene. Significantly, while Snrpn is widely expressed in adult tissues, U exon expression is detected only in postnatal brain from the paternal allele as well as in oocytes [15], [16], [18]. Expression of the U exons in oocytes correlates with imprint establishment in the female germ line, consistent with a role in establishing the maternal epigenotype.
As the 425Δ5-7A transgene displays appropriate imprinted expression and epigenetic imprinting marks, we hypothesized that U exon expression from this transgene would mimic the endogenous U exon expression patterns. To detect U exon transcripts arising exclusively from the transgene, we used a primer recognizing a loxP site that remained in the BAC after deletion of the region between Snrpn exons 5 and 7. We performed RT-PCR using a forward primer that anneals to both U1 and U2 (referred to as U-F1) and a reverse primer in the loxP sequence (Figure 4A). Replicating the endogenous locus, BAC transgene-encoded transcripts including the U exons were detected in P1 brain after paternal transmission of the transgene but not after maternal transmission (Figure S3). Importantly, these transcripts were also detected in transgenic ovary RNA from three-week-old females, corresponding with the timing of imprint establishment in oocytes (Figure 4B). Thus, U exon expression from the imprinted 425Δ5-7A transgene does mimic the endogenous expression pattern.

Snrpn U Exon Expression in the Germ Line Correlates with Maternal Imprint Establishment
We explored the role of Snrpn U exon transcription in directing imprinting at the PWS/AS locus by analyzing U exon expression in the developing germ line. Imprints are erased in the germ line in post-migratory germ cells between 10.5 to 12.5 days post coitus (dpc), yielding biallelic expression of Snrpn in 13.5 dpc primordial germ cells (PGCs) [27], [31]–[34]. Therefore, we examined 13.5 dpc PGCs for the presence of Snrpn U exon-containing transcripts, expecting a lack of expression since imprinting is not established until after birth in the maternal germ line. We utilized the U-F1 primer and a reverse primer complementary to Snrpn exon 3 to analyze expression in wild type, sex-segregated, purified 13.5 dpc PGC cDNAs (Figure 5A). Snrpn U exon-containing transcripts were not detected in either male or female 13.5 dpc PGCs (Figure 5B). RT-PCR was performed on the same cDNAs using the primer set spanning Snrpn exons 4 to 8 to demonstrate that Snrpn is expressed in both male and female 13.5 dpc PGCs. These data confirm that U exon transcription is not detectable in the germ line before imprints are established.

We further tested whether U exon expression contributed to imprinting by examining the postnatal ovary. Detecting expression in this tissue is vital as the DNA methylation imprint is established at the PWS-IC in growing oocytes. The Snrpn DMR methylation imprint first appears in growing oocytes between 5–25 days postpartum (dpp) [25], [26]. Therefore, we examined ovaries from wild type female mice to determine whether U exon transcription was active throughout this stage of imprint establishment. Ovaries obtained from females ranging in age from P1 to P20 were subjected RT-PCR analysis for transcripts spanning U-F1 to Snrpn exon 3. We detected U exon-containing transcripts in the ovary during the entire period of maternal imprint establishment at the PWS-IC (Figure 5C). This RT-PCR reaction revealed a larger amplicon than expected that was preferentially expressed in the developing ovaries. DNA sequencing of the PCR products indicated that the larger amplicon was derived from a transcript originating at U2 and included the Snrpn γ exon whereas the smaller product included only U1 and Snrpn exons 2 and 3.
Three Snrpn U Exons Constitute the AS-IC on the 425D18 BAC Transgene
We next sought to determine whether the Snrpn U exons are necessary for maternal imprinting of the 425Δ5-7 transgene. To do so, we used BAC recombineering to make targeted deletions of U1, U2, and U3, the three U exons contained within the 425Δ5-7 transgene. For each U exon, a 2.5 to 2.7 kb region spanning the exon was deleted to eliminate promoter function. This modified BAC was termed 425ΔU1-U3 (Figure 6A). Transgene copy number is an important element in imprinted transgene studies, especially as a multi-copy Snrpn transgene was previously shown to be imprinted while the same transgene in single copy was biparentally expressed [35]. We therefore limited our analysis to single copy 425ΔU1-U3 transgenic lines. In addition to providing the most stringent test of imprinting, single copy transgene lines eliminate uncertainty about which copy(s) of the transgene is being expressed and allow for accurate determination of DNA methylation status. The single copy line 425ΔU1-U3D was used for our initial analysis (Figure S1B). Unlike the 425Δ5-7 transgene, the 425ΔU1-U3D transgene was expressed in P1 brain upon both paternal and maternal inheritance, indicating an absence of AS-IC activity correlating with the deletion of the upstream exons (Figure 6B). We also examined the methylation imprint at the transgene Snrpn DMR and found that it was hypomethylated after both paternal and maternal transmission, demonstrating an absence of the maternal epigenetic imprint (Figure 6C). These experiments show that the 425Δ5-7 transgene loses its imprinted expression pattern as well as its epigenetic imprint upon removal of the three U exons contained within it, providing further evidence that these U exons play a major role in AS-IC function.

Imprinting Is Established in the Absence of the U Exons when Transcription through the PWS-IC Is Activated
We obtained one additional single copy line for the 425ΔU1-U3 transgene, 425ΔU1-U3H (Figure S1B). Unexpectedly, this transgene displayed imprinted expression of Snrpn (Figure 7B). We initially questioned whether critical sequences in the BAC had been rearranged upon integration but found that sequences for all single copy BAC transgenes were intact between U1 and the PWS-IC (Figure S4). Since the insertion sites for the transgenes in our study are unknown, we questioned whether transcripts initiated from a promoter upstream of the 425ΔU1-U3H transgene insertion site might proceed through the PWS-IC in the maternal germ line. We reasoned that this adventitious transcription could create an artificial imprint at the PWS-IC. To investigate this hypothesis, we performed RT-PCR on ovaries harvested from P15 females, including the imprinted 425ΔU1-U3H and 425Δ5-7A lines as well as the non-imprinted 425ΔU1-U3D line. As the 425ΔU1-U3 transgenes do not have the U exons intact, we used sequence from the Snrpn γ exon to generate our forward primer for RT-PCR since this exon is included in U exon transcripts generated during oocyte growth (Figure 5C). Unlike U1-U9, the Greek-lettered exons shown in Figure 1B have both splice acceptor signals in addition to splice donor signals [17]. We reasoned that the γ exon could be included in transcripts initiated from upstream of the transgene insertion site. As before, the reverse primer in the loxP sequence was used so as to avoid any signal from the endogenous allele. Figure 7C demonstrates that ovaries from the imprinted 425ΔU1-U3H line expressed Snrpn γ exon-containing transcripts that traversed the PWS-IC. These transcripts were also observed in the imprinted 425Δ5-7A line but not in the biparentally expressed 425ΔU1-U3D line. Thus, transcription through the PWS-IC in the ovary correlates with the establishment of the maternal imprint.

Discussion
Roughly 4% of individuals with AS have an imprinting defect, characterized by a paternal epigenotype at the PWS/AS locus on the maternally inherited chromosome. Of these individuals, 10–15% have a microdeletion that includes the AS-IC [36]. The AS-IC is thought to operate in the female germ line to epigenetically inactivate the PWS-IC and thereby silence the paternally expressed genes on the future maternal allele [9]. Among these maternally silenced genes is UBE3A-ATS, a transcript antisense to UBE3A. UBE3A-ATS is thought to silence UBE3A expression by an unknown mechanism [37]–[39]. In individuals lacking AS-IC function, UBE3A-ATS is expressed from the maternal allele in addition to the paternal allele and the consequent lack of UBE3A results in AS. Hypomethylation of the maternal SNRPN DMR is diagnostic of an AS imprinting defect [40].
The AS-IC functions in the female germ line, complicating investigations into imprinting mechanisms at the PWS/AS locus. Deletion analysis mapped the human AS-IC to 35 kb upstream of the PWS-IC but deletion of similarly positioned sequences in the mouse had no detectable consequence [19]. Here we used BAC transgenes to identify murine AS-IC activity. We found that 15 kb of sequence upstream of Snrpn exon 1 was insufficient to direct imprinted expression of Snrpn. Transgenes containing 100 kb of sequence 5′ to Snrpn exon 1 directed the appropriate imprinted expression of Snrpn. Furthermore, this sequence was sufficient to direct appropriate hypermethylation of the Snrpn DMR following maternal but not paternal transmission. We conclude that an AS-IC activity is contained within 15 to 100 kb of the PWS-IC. This region contains three alternative exons, at least two of which are expressed in oocytes and splice into Snrpn exon 2. Deletion of these U exons from the BAC transgene resulted in biparental Snrpn expression and DNA hypomethylation at the PWS-IC, characteristic of an imprinting defect. Surprisingly, a second single copy line lacking the U exons displayed appropriate maternal silencing. In contrast to the biparentally expressed transgene but similar to imprinted transgenes with the U exons intact, this line exhibited transcription through the PWS-IC, consistent with the notion that transcription is an essential element of AS-IC activity. Lastly, we demonstrated that U exon transcription is undetectable in fetal oogonia as they are entering meiotic prophase, a time when imprinting is absent, but becomes evident after birth and prior to the application of the DNA methylation imprint at the PWS-IC. We conclude that the U exons exhibit AS-IC activity as previously proposed [8], [15].
Snrpn transcription becomes biallelic in PGCs as they colonize the gonad at midgestation [31]. This stage of imprinting erasure in the germ line coincides with the removal of the DNA methylation imprint at the PWS-IC [27]. The appearance of the DNA methylation imprint is tied to postnatal oocyte growth [25], [26]. Our data support a model in which growing oocytes direct transcription from the U exons through the PWS-IC. This transcription is necessary to epigenetically modify the PWS-IC leading to the maternal imprint (Figure 8). Importantly, our results do not distinguish whether the RNA transcript or the act of transcription is essential for AS-IC activity. However, since transcription presumably arising from sequences flanking the BAC insertion site appears to be sufficient for imprinting in the 425ΔU1-U3H line, we suggest that the U exon sequences per se are not required.
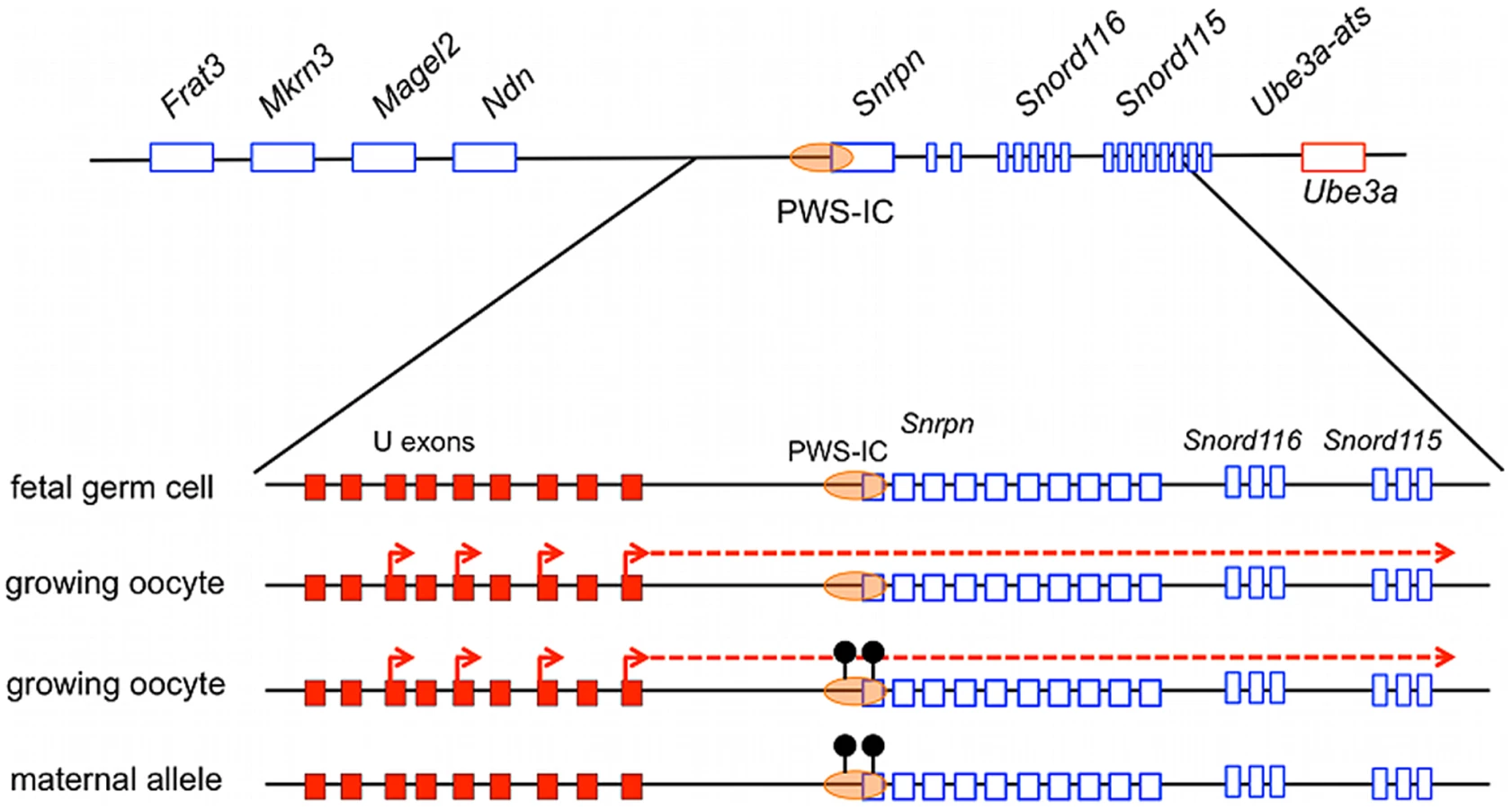
Our data support a model in which U exon promoter activity supplies AS-IC function in the mouse. The human AS-IC, however, includes two upstream exons that are internal to transcripts encoded in brain RNA. If the human AS-IC does not possess promoter activity in oocytes, we speculate that the AS-IC may serve an RNA antitermination or stabilization role.
In both human and mouse, splicing of the upstream exons to Snrpn exon 2 can encode the Snurf polypeptide [17], [41]. It is unknown whether Snurf is expressed in oocytes but the absence of an overt phenotype in Snurf knockout mice indicates that it is not involved in the imprinting process [42].
Our BAC transgene system allows simultaneous assessment of both the transgene and endogenous locus. U exon transcription arising from the endogenous locus does not impose imprinting upon a maternally transmitted transgene harboring a PWS-IC but lacking U exons (Figure 2 and Figure 6B). This observation suggests that if the RNA transcript is an essential element of the imprinting machinery, then it does not function in trans.
Wu et al. [20] described two mutants with a maternal imprinting defect. Maternal transmission of an 80 kb deletion spanning from 10 to 90 kb upstream of Snrpn exon 1 resulted in an extensive loss of DNA methylation at the PWS-IC in only three of eight offspring. This deletion removes only U1 and U3, leaving the remaining seven U exons intact. Our model suggests that extensive imprinting defects were not observed in the remaining offspring due to compensatory transcription arising from the remaining U exons. The second mutant described by these investigators exhibited a highly penetrant imprinting defect leading to expression of paternal genes from the maternal allele and reduced expression of the Ube3a gene product, E6AP. This mutant carried an insertion 10 kb upstream of Snrpn exon 1 that was comprised of a puromycin resistance cassette in the opposite transcriptional orientation to Snrpn, exons 3–9 of an Hprt minigene, and a duplication of 6 kb of endogenous sequence. Our findings support a model in which the insertion interferes with transcription from the U exons in oocytes.
The imprinting activity of the human AS-IC element has been tested in mice. Five independent lines of a human P1 phage transgene that included both the human AS-IC and PWS-IC failed to show evidence of imprinted gene expression in the brain [35]. This lack of imprinting may be explained by our more recent observation that a human PWS-IC substituted for the endogenous murine PWS-IC can acquire a DNA methylation imprint in oocytes but this imprint is subsequently lost during development [38]. Shemer et al. [10] characterized transgenes containing the human AS-IC linked to the murine PWS-IC and observed preferential Snrpn expression following paternal inheritance. Transcription through the PWS-IC during oogenesis was not examined in these transgenes but our model suggests that the partial imprinting seen in these transgenes resulted from transcription of the human upstream exons.
Zogel et al. [43] reported that a maternally inherited polymorphism at the AS-IC increases the risk of imprinting defects leading to AS. Our observations strongly support these investigators′ hypothesis that this polymorphism may affect a transcription factor recognition site. Our model suggests that this polymorphism affects transcription through the PWS-IC during oogenesis, decreasing the efficiency of epigenetic modification at the PWS-IC and thereby raising the risk of AS imprinting defects.
Transcription is implicated in allelic silencing at several imprinted gene clusters (See Peters and Robson [44] for a review). Our results at the PWS/AS locus are consistent with the observation that in growing oocytes, transcripts traverse maternal germ line DMRs at eight locations, suggesting that transcription is commonly associated with the establishment of maternal imprints [45]. A recent report indicates that 35% of CpG islands that are methylated in oocytes are downstream of active promoters [46]. The role of transcription through a germ line DMR in establishing a DNA methylation imprint in oocytes has been tested at the Gnas locus [45]. At this locus, Nesp transcription is active in oocytes and traverses two maternal DMRs. Similar to the U transcripts at the PWS/AS locus, appearance of the Nesp transcript at the Gnas locus temporally precedes the establishment of the maternal methylation imprint in growing oocytes. Furthermore, truncation of the Nesp transcript led to a loss of the DNA methylation imprint at several maternal DMRs within the Gnas locus and activation of normally silent maternal transcripts. Chotalia et al. [45] suggested several mechanisms by which transcription might contribute to DNA methylation including an interaction of the transcription complex with the methylation machinery, creating an open chromatin environment allowing DNA methylation machinery access to the locus, or recruiting factors necessary for epigenetic modification. These same mechanisms could operate to set the methylation imprint at the PWS/AS locus. Similar to the Gnas locus, truncation of the U exon transcripts upstream of the PWS-IC may confirm their role in establishing imprinting throughout the PWS/AS locus.
Materials and Methods
Ethics Statement
All animal procedures in this work were reviewed and approved by the University of Florida Institutional Animal Care and Use Committee.
BAC DNAs
BACs 380J10 and 425D18 were originally identified by application of a Snrpn probe to the RPCI-23 library prepared from C57BL/6 DNA (Research Genetics). BAC 380J10 is 232.3 kb and contains chromosome 7 nucleotides (nt) 67165323 to 66935240. This region includes 15.3 kb upstream of the major Snrpn transcription start site to 192.1 kb 3′ sequence containing nearly the entire complement of Snord116 repeats. BAC 425D18 is 192.3 kb and contains sequences from nt 67250191 to 67057909. This span includes 100.1 kb upstream of the major Snrpn transcription start site and 69.5 kb sequence 3′ to the Snrpn polyadenylation site.
Recombineering procedures were performed in bacterial strains DY380, EL250 and EL350 as described [47]. First, a loxP site in the BACe3.6 vector was replaced with a blasticidin resistance cassette. To create a reporter gene in the BAC, a deletion spanning Snrpn exons 5 to 7 was engineered by homologous recombination with a floxed kanamycin (kan) resistance cassette followed by Cre-mediated removal of the kan resistance cassette in EL350 cells. This deletion removes sequences previously shown to be unimportant for imprinting [21]. The three U exons in this modified BAC were deleted individually by first subcloning a 4.6 to 5.0 kb region surrounding each U exon and subsequently inserting a kan resistance cassette into each clone. These clones were then used to recombineer the modified 425D18 BAC. The U1 deletion removes 984 bp 5′ to 1576 bp 3′ of U1. The U2 deletion removes sequences from 1032 bp 5′ to 1296 bp 3′ of U2. The U3 deletion removes sequences from 834 bp 5′ to 1575 bp 3′ of U3. In each case, the kan resistance cassette was removed by recombineering. Recombination between lox sites at different U exons was prevented by using kan resistance cassettes flanked by FRT sites at U1 and by incompatible lox sequences at U2 and U3 [48]. Transfer of each deletion to the BAC was confirmed by Southern blot analysis and DNA sequencing.
Transgenic Mice
C57BL/6 BAC DNA was purified by CsCl gradient centrifugation and supercoiled DNA was injected into fertilized FVB/N oocytes. Strains were maintained on an FVB background except where noted for the DNA methylation studies.
Southern Blot Analysis
Genomic DNA was digested with restriction endonucleases as indicated and analyzed by Southern blot. The 380J10 copy number blot was probed with a 770 bp EcoRI fragment approximately 7.8 kb upstream of Snrpn exon 1. The 425D18 copy number blot was probed with a 291 bp EcoRI fragment located 5′ to the deletion between Snrpn exons 5 and 7. Relative band intensity was digitized on the Storm 860 PhosphorImager (GE Healthcare) and analyzed with Image Quant TL software for transgene copy number analyses. Probes for Southern blots demonstrating the integrity of the 425D18 derived BAC transgenes were generated from PCR products (H, E,L) (see Dataset S1 for primer sequences) or restriction fragments (304 : 800 bp PstI fragment, 210 : 770 bp EcoRI fragment).
Analysis of Gene Expression
RNA was isolated from the indicated tissues by homogenization with RNA-Bee reagent (Tel-Test, Inc.) following manufacturer's instructions. RNA was subject to Northern blot analysis or DNase treated and subsequently reverse-transcribed with Superscript II (Invitrogen). PCR reactions were performed under standard conditions. Primer sequences listed in Dataset S1. The Snrpn Northern blot probe was generated from a 480 bp ClaI-PstI fragment.
DNA Methylation Analysis
Bisulfite sequence analysis was performed on whole brain genomic DNA. DNA was subject to bisulfite conversion as previously described [49]. PCR amplification of the Snrpn DMR was performed on the bisulfite converted DNA using the primer set W18-F/W19-R [19] (Dataset S1). An initial 15 minute denaturation step was followed by 38 cycles of 94°C for 45 s, 54°C for 60 s and 72°C for 90 s with a 10 minute final extension at 72°C. Amplified products were cloned into the pGEM-T Easy Vector System (Promega). Plasmid sequencing was performed with ABI Prism BigDye terminator (PerkinElmer) at the UF Center for Epigenetics.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. LewisAReikW 2006 How imprinting centres work. Cytogenet Genome Res 113 81 89
2. RazinACedarH 1994 DNA Methylation and Genomic Imprinting. Cell 77 473 476
3. MargueronRTrojerPReinbergD 2005 The key to development: interpreting the histone code? Curr Opin Genet Dev 15 163 176
4. KitsbergDSeligSBrandeisMSimonIKeshetI 1993 Allele-specific replication timing of imprinted gene regions. Nature 364 459 463
5. KishinoTLalandeMWagstaffJ 1997 UBE3A/E6-AP mutations cause Angelman syndrome. Nature Genetics 15 70 73
6. MatsuuraTSutcliffeJSFangPGaljaardR-JJiangY-H 1997 De novo truncating mutations in E6-AP ubiquitin-protein ligase gene (UBE3A) in Angelman syndrome. Nature Genetics 15 74 77
7. BuitingKSaitohSGrossSDittrichBSchwartzS 1995 Inherited microdeletions in the Angelman and Prader-Willi syndromes define an imprinting centre on human chromosome 15. Nat Genet 9 395 400
8. DittrichBBuitingKKornBRickardSBuxtonJ 1996 Imprint switching on human chromosome 15 may involve alternative transcripts of the SNRPN gene. Nat Genet 14 163 170
9. BrannanCIBartolomeiMS 1999 Mechanisms of genomic imprinting. Current Opinion in Genetics and Development 9 164 170
10. ShemerRHershkoAYPerkJMostoslavskyRTsuberiB 2000 The imprinting box of the Prader-Willi/Angelman syndrome domain. Nature Genetics 26 440 443
11. BuitingKLichCCottrellSBarnicoatAHorsthemkeB 1999 A 5-kb imprinting center deletion in a family with Angelman syndrome reduces the shortest region of deletion overlap to 880 bp. Human Genetics 105 665 666
12. OhtaTGrayTARoganPKBuitingKGabrielJM 1999 Imprinting-mutation mechanisms in Prader-Willi syndrome. American Journal of Human Genetics 64 397 413
13. FarberCDittrichBBuitingKHorsthemkeB 1999 The chromosome 15 imprinting centre (IC) region has undergone multiple duplication events and contains an upstream exon of SNRPN that is deleted in all Angelman syndrome patients with an IC microdeletion. Hum Mol Genet 8 337 343
14. WawrzikMSpiessANHerrmannRBuitingKHorsthemkeB 2009 Expression of SNURF-SNRPN upstream transcripts and epigenetic regulatory genes during human spermatogenesis. Eur J Hum Genet 17 1463 1470
15. MapendanoCKKishinoTMiyazakiKKondoSYoshiuraK 2006 Expression of the Snurf-Snrpn IC transcript in the oocyte and its putative role in the imprinting establishment of the mouse 7C imprinting domain. J Hum Genet 51 236 243
16. BresslerJTsaiTFWuMYTsaiSFRamirezMA 2001 The SNRPN promoter is not required for genomic imprinting of the Prader - Willi/Angelman domain in mice. Nature Genetics 28 232 240
17. LandersMBancescuDLLe MeurERougeulleCGlatt-DeeleyH 2004 Regulation of the large (∼1000 kb) imprinted murine Ube3a antisense transcript by alternative exons upstream of Snurf/Snrpn. Nucleic Acids Res 32 3480 3492
18. Le MeurEWatrinFLandersMSturnyRLalandeM 2005 Dynamic developmental regulation of the large non-coding RNA associated with the mouse 7C imprinted chromosomal region. Dev Biol 286 587 600
19. PeeryEGElmoreMDResnickJLBrannanCIJohnstoneKA 2007 A targeted deletion upstream of Snrpn does not result in an imprinting defect. Mamm Genome 18 255 262
20. WuMYChenKSBresslerJHouATsaiTF 2006 Mouse imprinting defect mutations that model Angelman syndrome. Genesis 44 12 22
21. YangTAdamsonTEResnickJLLeffSWevrickR 1998 A mouse model for Prader-Willi syndrome imprinting-centre mutations. Nature Genetics 19 25 31
22. SutcliffeJSNakaoMChristianSOrstavikKHTommerupN 1994 Deletions of a differentially methylated CpG island at the SNRPN gene define a putative imprinting control region. Nat Genet 8 52 58
23. GabrielJMGrayTAStubbsLSaitohSOhtaT 1998 Structure and function correlations at the imprinted mouse Snrpn locus. Mamm Genome 9 788 793
24. ShemerRBirgerYRiggsADRazinA 1997 Structure of the imprinted mouse Snrpn gene and establishment of its parental-specific methylation pattern. Proc Natl Acad Sci U S A 94 10267 10272
25. HiuraHObataYKomiyamaJShiraiMKonoT 2006 Oocyte growth-dependent progression of maternal imprinting in mice. Genes Cells 11 353 361
26. LuciferoDMannMRBartolomeiMSTraslerJM 2004 Gene-specific timing and epigenetic memory in oocyte imprinting. Hum Mol Genet 13 839 849
27. HajkovaPErhardtSLaneNHaafTEl-MaarriO 2002 Epigenetic reprogramming in mouse primordial germ cells. Mech Dev 117 15 23
28. WakelandEKMorelLAcheyKYuiMLongmateJ 1997 Speed congenics: A classic technique in the fast lane (relatively speaking). Immunology Today 18 472 477
29. OhtaTBuitingKKokkonenHMcCandlessSHeegerS 1999 Molecular mechanism of angelman syndrome in two large families involves an imprinting mutation. Am J Hum Genet 64 385 396
30. SaitohSBuitingKRoganPKBuxtonJLDriscollDJ 1996 Minimal definition of the imprinting center and fixation of chromosome 15q11–q13 epigenotype by imprinting mutations. Proc Natl Acad Sci U S A 93 7811 7815
31. SzaboPEMannJR 1995 Biallelic expression of imprinted genes in the mouse germ line: implications for erasure, establishment, and mechanisms of genomic imprinting. Genes Dev 9 1857 1868
32. SzaboPEHubnerKScholerHMannJR 2002 Allele-specific expression of imprinted genes in mouse migratory primordial germ cells. Mech Dev 115 157 160
33. ObataYKonoT 2002 Maternal Primary Imprinting Is Established at a Specific Time for Each Gene throughout Oocyte Growth. J Biol Chem 277 5285 5289
34. YamazakiYMannMRLeeSSMarhJMcCarreyJR 2003 Reprogramming of primordial germ cells begins before migration into the genital ridge, making these cells inadequate donors for reproductive cloning. Proc Natl Acad Sci U S A 100 12207 12212
35. BlaydesSMElmoreMYangTBrannanCI 1999 Analysis of murine Snrpn and human SNRPN gene imprinting in transgenic mice. Mammalian Genome 10 549 555
36. BuitingK 2010 Prader-Willi syndrome and Angelman syndrome. Am J Med Genet C Semin Med Genet 154C 365 376
37. ChamberlainSJBrannanCI 2001 The Prader-Willi syndrome imprinting center activates the paternally expressed murine Ube3a antisense transcript but represses paternal Ube3a. Genomics 73 316 322
38. JohnstoneKADuBoseAJFuttnerCRElmoreMDBrannanCI 2006 A human imprinting centre demonstrates conserved acquisition but diverged maintenance of imprinting in a mouse model for Angelman syndrome imprinting defects. Hum Mol Genet 15 393 404
39. RougeulleCCardosoCFontesMColleauxLLalandeM 1998 An imprinted antisense RNA overlaps UBE3A and a second maternally expressed transcript. Nat Genet 19 15 16
40. GlennCCSaitohSJongMTFilbrandtMMSurtiU 1996 Gene structure, DNA methylation, and imprinted expression of the human SNRPN gene. Am J Hum Genet 58 335 346
41. GrayTASaitohSNichollsRD 1999 An imprinted, mammalian bicistronic transcript encodes two independent proteins. Proc Natl Acad Sci U S A 96 5616 5621
42. TsaiTFJiangYHBresslerJArmstrongDBeaudetAL 1999 Paternal deletion from Snrpn to Ube3a in the mouse causes hypotonia, growth retardation and partial lethality and provides evidence for a gene contributing to Prader-Willi syndrome. Human Molecular Genetics 8 1357 1364
43. ZogelCBohringerSGrossSVaronRBuitingK 2006 Identification of cis - and trans-acting factors possibly modifying the risk of epimutations on chromosome 15. Eur J Hum Genet 14 752 758
44. PetersJRobsonJE 2008 Imprinted noncoding RNAs. Mamm Genome 19 493 502
45. ChotaliaMSmallwoodSARufNDawsonCLuciferoD 2009 Transcription is required for establishment of germline methylation marks at imprinted genes. Genes Dev 23 105 117
46. SmallwoodSATomizawaSKruegerFRufNCarliN 2011 Dynamic CpG island methylation landscape in oocytes and preimplantation embryos. Nat Genet 43 811 814
47. LeeECYuDMartinez de VelascoJTessarolloLSwingDA 2001 A highly efficient Escherichia coli-based chromosome engineering system adapted for recombinogenic targeting and subcloning of BAC DNA. Genomics 73 56 65
48. SiegelRWJainRBradburyA 2001 Using an in vivo phagemid system to identify non-compatible loxP sequences. FEBS Lett 505 467 473
49. ClarkSJHarrisonJPaulCLFrommerM 1994 High sensitivity mapping of methylated cytosines. Nucleic Acids Res 22 2990 2997
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 12
Nejčtenější v tomto čísle
- Targeted Proteolysis of Plectin Isoform 1a Accounts for Hemidesmosome Dysfunction in Mice Mimicking the Dominant Skin Blistering Disease EBS-Ogna
- The RNA Silencing Enzyme RNA Polymerase V Is Required for Plant Immunity
- The FGFR4-G388R Polymorphism Promotes Mitochondrial STAT3 Serine Phosphorylation to Facilitate Pituitary Growth Hormone Cell Tumorigenesis
- Target Site Recognition by a Diversity-Generating Retroelement