
Multiple Roles and Interactions of and in Development of the Respiratory System
Normal development of the respiratory system is essential for survival and is regulated by multiple genes and signaling pathways. Both Tbx4 and Tbx5 are expressed throughout the mesenchyme of the developing lung and trachea; and, although multiple genes are known to be required in the epithelium, only Fgfs have been well studied in the mesenchyme. In this study, we investigated the roles of Tbx4 and Tbx5 in lung and trachea development using conditional mutant alleles and two different Cre recombinase transgenic lines. Loss of Tbx5 leads to a unilateral loss of lung bud specification and absence of tracheal specification in organ culture. Mutants deficient in Tbx4 and Tbx5 show severely reduced lung branching at mid-gestation. Concordant with this defect, the expression of mesenchymal markers Wnt2 and Fgf10, as well as Fgf10 target genes Bmp4 and Spry2, in the epithelium is downregulated. Lung branching undergoes arrest ex vivo when Tbx4 and Tbx5 are both completely lacking. Lung-specific Tbx4 heterozygous;Tbx5 conditional null mice die soon after birth due to respiratory distress. These pups have small lungs and show severe disruptions in tracheal/bronchial cartilage rings. Sox9, a master regulator of cartilage formation, is expressed in the trachea; but mesenchymal cells fail to condense and consequently do not develop cartilage normally at birth. Tbx4;Tbx5 double heterozygous mutants show decreased lung branching and fewer tracheal cartilage rings, suggesting a genetic interaction. Finally, we show that Tbx4 and Tbx5 interact with Fgf10 during the process of lung growth and branching but not during tracheal/bronchial cartilage development.
Published in the journal:
. PLoS Genet 8(8): e32767. doi:10.1371/journal.pgen.1002866
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002866
Summary
Normal development of the respiratory system is essential for survival and is regulated by multiple genes and signaling pathways. Both Tbx4 and Tbx5 are expressed throughout the mesenchyme of the developing lung and trachea; and, although multiple genes are known to be required in the epithelium, only Fgfs have been well studied in the mesenchyme. In this study, we investigated the roles of Tbx4 and Tbx5 in lung and trachea development using conditional mutant alleles and two different Cre recombinase transgenic lines. Loss of Tbx5 leads to a unilateral loss of lung bud specification and absence of tracheal specification in organ culture. Mutants deficient in Tbx4 and Tbx5 show severely reduced lung branching at mid-gestation. Concordant with this defect, the expression of mesenchymal markers Wnt2 and Fgf10, as well as Fgf10 target genes Bmp4 and Spry2, in the epithelium is downregulated. Lung branching undergoes arrest ex vivo when Tbx4 and Tbx5 are both completely lacking. Lung-specific Tbx4 heterozygous;Tbx5 conditional null mice die soon after birth due to respiratory distress. These pups have small lungs and show severe disruptions in tracheal/bronchial cartilage rings. Sox9, a master regulator of cartilage formation, is expressed in the trachea; but mesenchymal cells fail to condense and consequently do not develop cartilage normally at birth. Tbx4;Tbx5 double heterozygous mutants show decreased lung branching and fewer tracheal cartilage rings, suggesting a genetic interaction. Finally, we show that Tbx4 and Tbx5 interact with Fgf10 during the process of lung growth and branching but not during tracheal/bronchial cartilage development.
Introduction
The development of the respiratory system represents an evolutionary hallmark that allowed vertebrates to survive on land utilizing air as a source of oxygen. Because the respiratory system is dispensable for embryonic survival in mammals, defects in development of the respiratory system manifest at or after birth. Indeed, abnormal development of the respiratory system in humans is associated with multiple disorders such as tracheal/bronchial atresia, tracheoesophageal fistula, bronchogenic cysts, pulmonary/lobar atresia and pulmonary hypoplasia [1]. Thus, it is important to understand the genetic basis of development of the respiratory system.
In the mouse embryo, the endodermal foregut tube is patterned by signals from the lateral plate mesoderm leading to specification of the lung and trachea at embryonic day (E) 9.0 (19–24 somites). Nkx2.1 has been identified as the earliest marker of lung endoderm specification. At E9.25 (25–28 somites), the primary lung buds appear as ventro-lateral outpouchings of the foregut connected ventrally by the tracheal primordium. The lung buds grow in a ventro-posterior direction and continue to elongate until E11.5. The point of connection of the lung buds is thought to be the origin of the tracheal tube, which separates from the esophagus in a caudal to cranial direction by E11.5 [2], [3]. In the mouse, the left lung bud remains a single lobe and the right lung bud forms 4 lobes - cranial, medial, caudal and accessory [2]. The airways undergo a stereotypic pattern of branching beginning at E11.5 [4]; development and maturation of the alveoli occurs later.
Genes involved in different signaling pathways, including Wnt2, Fgf10, Bmp4, Shh and retinoic acid synthesis genes, have been shown to play important roles in lung specification and branch formation. Complete absence of both Wnt2 and Wnt2b in mesenchyme surrounding the anterior foregut or absence of β-catenin in the foregut epithelium leads to a loss of specification of lung primordia as seen by the absence of Nkx2.1 expression [5]–[7]. Embryos lacking Fgf10, which is normally expressed in mesenchyme surrounding the epithelial branching tips, form a short trachea but have no lungs [8], [9]. Inhibition of epithelial Bmp4 signaling by overexpression of Xnoggin leads to a decrease in lung size and irregularly shaped lung lobes [10]. Shh null mutant mice have only a rudimentary lung sac due to branching severe branching defect [11]. Additionally, conditional inactivation of Shh in lung epithelial cells leads to the formation of hypoplastic lungs with reduced branching of the peripheral tubules [12]. Retinoic acid receptor (RAR) α and RAR β2 double null mutants show left lung agenesis and a hypoplastic right lung at E18.5 [13]. Thus, genes expressed in both the mesenchyme and the epithelium are essential for correct lung bud specification and branching.
After E11.5, mesenchyme surrounding the dorsal aspect of the trachea differentiates into the trachealis smooth muscle. Mesenchyme surrounding the ventral aspect of the trachea and lateral aspect of the main stem bronchi segments and differentiates into C shaped rings composed of chondrocytes. Ventral tracheal cartilage is formed by migration of cells that undergo mesenchymal condensation [14]. Sox9 has been implicated as an important regulator of mesenchymal condensation and chondrocyte differentiation [15], [16]. In chondrocyte cultures it has been shown that in addition to Sox9, FGF2, Igf1, Tgfβ2 and Bmp2 enhance chondrocyte formation [14], [17]. Mutations in a number of genes including Shh, Sox2, retinoic acid synthesis genes and Fgf signaling pathway genes have been shown to affect cartilage ring formation [11]–[13], [18]–[21]. Fgf10 mutants form a partial tracheal tube in spite of the failure of lung formation [8], [9]. Recent evidence shows that loss of Fgf10 leads to defects in tracheal ring formation and that overexpression of Fgf10 between E11.5 and E13.5 disrupts tracheal rings by altering the periodic expression of Shh in the trachea [22].
The T-box transcription factor genes are important during embryonic development. All members of this gene family contain a conserved DNA-binding T-box domain, which binds to a conserved sequence, the T-box binding element, to activate or repress transcription of specific target genes [23]. All Tbx2 subfamily genes, Tbx2, Tbx3, Tbx4 and Tbx5 are expressed in the developing chick lung buds and trachea between stages 15–21 [24]. In the mouse, Tbx1 is expressed in lung epithelium at E12.5, Tbx2 and Tbx3 are expressed in lung mesenchyme at E11.5, and Tbx4 and Tbx5 are expressed in both lung and trachea mesenchyme at E12.5 and later [25]. Tbx1 homozygous null mutants die at birth due to severe heart defects; the lungs are never fully inflated [26] but lung development has not been further investigated. In Tbx4 homozygous mutants, lung buds form but the embryos die at E10.5 due to failure of allantois development and the subsequent lack of chorio-allantoic fusion leading to placental insufficiency [27]. Tbx5 mutants die around E10 due to defects in heart development [28]; lung development has not been previously investigated. Antisense oligonucleotide depletion of both Tbx4 and Tbx5, but not Tbx2 and Tbx3, in lung organ cultures results in inhibition of branching and loss of Fgf10 expression [29] suggesting a role for these factors in lung branching. In the chick embryo, interference with Tbx4 function leads to a reduction in Fgf10 expression in lung mesenchyme and inhibits lung bud formation. Ectopic expression of Tbx4 leads to ectopic expression of Fgf10 and Nkx2.1 and lung bud formation in the esophagus. Additionally, ectopic expression of Tbx4 at the boundary between the trachea and the esophagus can lead to lack of separation of these two structures, resulting in a tracheoesophageal fistula [30]. In humans, Tbx5 mutations cause Holt Oram syndrome characterized by heart and forelimb abnormalities. A single de-novo mutation in TBX5 has been linked to right lung agenesis [31].
To study the roles of Tbx4 and Tbx5 in lung and trachea development in the mouse, we made use of conditional alleles to bypass early embryonic lethality. We studied three distinct processes, namely 1) lung bud and trachea specification, 2) lung branching morphogenesis, and 3) tracheal/bronchial cartilage formation. We show that during early stages of development, Tbx5 is important for specification of the lung buds and the trachea. After specification, Tbx4 and Tbx5 interact during lung growth and branching and the regulation of branching is dependent on Fgf10 signaling. Additionally, Tbx4 and Tbx5 interact in the formation of mesenchymal condensations, which ultimately form the tracheal/bronchial cartilage rings independent of Fgf10 signaling.
Results
Expression of Tbx4 and Tbx5 in the developing lung and trachea
Tbx5 expression is first detected using in situ hybridization (ISH) at E9.0 (24 somites) in the mesenchyme of the lung and trachea primordia, concurrent with Nkx2.1 expression in the ventral foregut epithelium (Figure 1A–1D). The anterior extent of expression of both genes coincides with the posterior extent of the third pharyngeal pouch (red arrow in Figure 1A, 1B). Tbx4 expression is detected in the lung buds when they first appear a few hours later at E9.25 (28 somites) in a pattern similar to Tbx5 (Figure 1E, 1F). Tbx4 and Tbx5 are expressed at E11.5, E13.5 and E15.5 throughout the lung mesenchyme but not in the epithelium (Figure 1G–1L) [25].
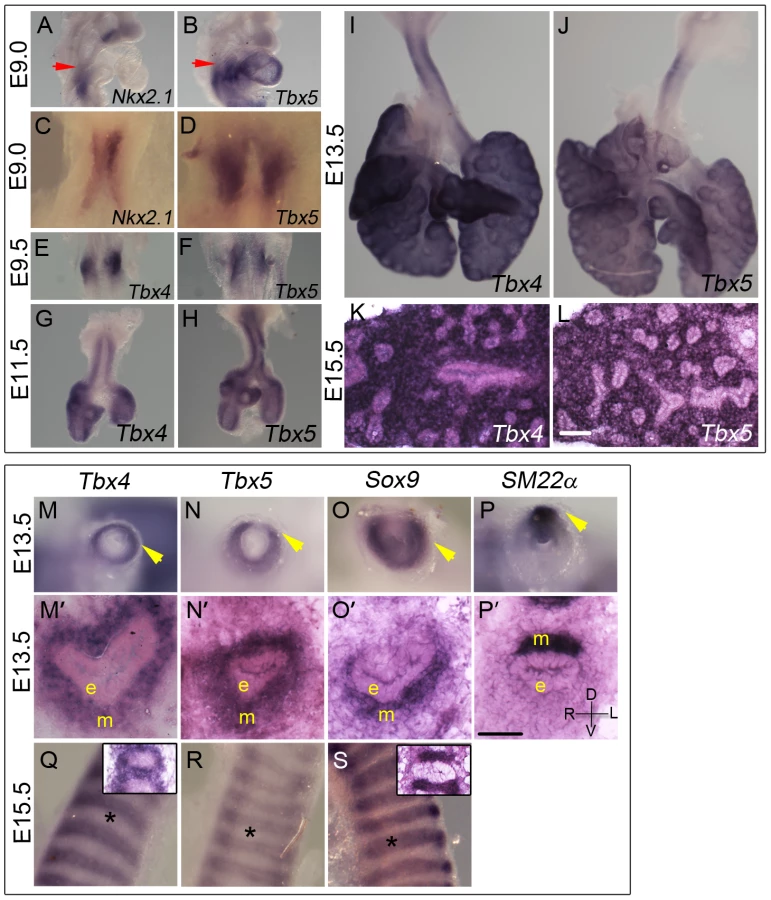
Both Tbx4 and Tbx5 show a dynamic expression pattern in the developing trachea. At E11.5 and E13.5 both genes are expressed throughout the tracheal mesenchyme surrounding the epithelium but are excluded from the epithelium and the outermost layer, the mesothelium (Figure 1G, 1H, 1M, 1M′, 1N, 1N′). At these stages, Tbx4 and Tbx5 are expressed in ventral tracheal cells that also express Sox9 (Figure 1O, 1O′) [32] and in dorsal tracheal cells that also express SM22α (Figure 1P, 1P′) [33]. Within the ventral mesenchyme at E15.5, Tbx4 and Tbx5 expression is restricted to mesenchyme between and surrounding cartilage condensations (Figure 1Q, 1R) in a pattern complementary to that of Sox9, which is restricted to condensing mesenchyme of the cartilage rings at this stage (Figure 1S).
Efficiency of recombination of conditional alleles
The genotypes of embryos used in this study and the corresponding descriptive shorthand nomenclature are shown in Table 1. PCR genotyping was used to determine the efficiency of recombination of the conditional alleles. For embryos carrying the tamoxifen-inducible CreER transgene, a dose of 8 mg tamoxifen was injected into pregnant females at E9.0 and embryos were dissected at E12.5. Yolk sacs were analyzed as an estimate of recombination in the whole embryo. Both alleles of Tbx4fl/fl embryos were completely recombined at E12.5 to produce the mutant allele at this dose of tamoxifen (Figure S1A), but the single floxed allele of Tbx5fl/+ embryos was only partially recombined to the mutant form (Figure S1E). Doses of tamoxifen higher than 8 mg at E9.0 lead to a loss of pregnancy. When 7 mg tamoxifen was injected at E8.5, complete recombination of the Tbx4fl alleles (Figure S1B) and the single Tbx5fl allele (Figure S1F) was obtained at E13.5.

In lung bud cultures a concentration of 1 µM 4-OH tamoxifen produced near-complete recombination of the Tbx4fl allele after 24 hours (Figure S1C) whereas the Tbx5fl allele was only partially recombined (Figure S1G). Virtually complete recombination of all floxed alleles was achieved after 4 days of culture (Figure S1D, S1H). These data suggest that the Tbx5fl allele has a lower efficiency of Cre-mediated recombination than the Tbx4fl allele. Thus we assume that there may be some residual Tbx5 activity from the Tbx5fl allele in the in vivo experiments, even in the presence of the CreER and Tbx4cre alleles.
Early loss of Tbx5, but not Tbx4, leads to a unilateral loss of lung bud specification and absence of tracheal specification
To explore the role of Tbx4 and Tbx5 in the earliest stages of lung and trachea specification, foregut culture [34] was used with Nkx2.1 as a marker of specification. This ex vivo technique allows for analysis of mutants in culture, circumventing early embryonic lethality of the Tbx4 homozygous mutants due to allantois defects and Tbx5 homozygous mutants due to heart defects. When foreguts and surrounding tissue are isolated at E8.75 (8–16 somites), the foregut tube is devoid of Nkx2.1 expression and the lung buds are not present [6]. At the end of 3 or 4 days of culture lung buds and trachea have formed as seen by Nkx2.1 expression (Figure 2A, 2D, 2G). Nkx2.1 is also expressed in the thyroid primordia at this stage (Figure 2D–2I and [35]). Expression of Tbx4 and Tbx5 was confirmed in control foreguts that were cultured for 4 days (Figure 2B, 2C). Foreguts from E8.5 embryos were cultured in the presence of 4-hydroxy (OH) tamoxifen and analyzed for Nkx2.1 expression. Reduction of Tbx5 alone lead to a lack of Nkx2.1 expression in one of the lung buds after 3 or 4 days of culture (Figure 2E and 2H, respectively) suggesting a unilateral loss of lung bud specification. Removal of Tbx4 alone did not affect Nkx2.1 expression after 3 days of culture (data not shown) and removal of Tbx4 in addition to Tbx5 did not exacerbate the Tbx5 phenotype (Figure 2F, 2I). Therefore, Tbx5 but not Tbx4 is important for the bilateral specification of lung buds ex vivo. Wnt2 and Wnt2b, genes essential for specification of respiratory primordia [5], were analyzed in the conditional Tbx5 null foreguts. Wnt2 expression was reduced (Figure 2J, 2K) and Wnt2b expression was absent (Figure 2L, 2M) suggesting that Tbx5 lies upstream of these genes in regulating the process of specification.

With respect to tracheal specification, Nkx2.1 expression was not observed in the foregut tube after 3 or 4 days of culture in embryos lacking Tbx5 (arrowheads in Figure 2E, 2H) suggesting a lack of tracheal specification in the absence of Tbx5. Additional loss of Tbx4 alleles did not alter the phenotype (Figure 2F, 2I), supporting a role for Tbx5 in the specification of the trachea, independent of Tbx4.
Lung branching is severely affected by the loss of Tbx4 and Tbx5
To analyze the effect of loss of Tbx4 and Tbx5 on lung branching in vivo we made use of the tamoxifen-inducible CreER transgene. Tamoxifen was injected at E8.75 (8–16 somites), late enough to bypass lethality but well before lung branching begins. Embryos with CreER and different combinations of the Tbx4fl and Tbx5fl alleles were examined at E12.5 and E13.5. Conditional Tbx4 null lungs were similar to controls at E12.5 but had fewer branching tips at E13.5 (Figure 3A, 3B, 3E, 3F). Conditional Tbx4;Tbx5 double heterozygous lungs (Figure 3C) were smaller in size and had fewer branching tips than conditional Tbx4 null lungs at E13.5 (Figure 3B) but were more advanced developmentally (Figure 3E, 3F) than conditional Tbx4 null;Tbx5 heterozygous lungs (Figure 3D), which were severely retarded. Lobation in the right lung was disrupted in conditional Tbx4 null;Tbx5 heterozygous lungs: the accessory lobe was missing and only rudimentary cranial and medial lobes were present (Figure 3H). The lobes had a fused appearance (Figure 3H′) suggesting a failure of separation. Histologically, there were no obvious structural defects other than an overall reduction in size (Figure 3G, 3G′, 3H, 3H′).

Conditional Tbx4 null;Tbx5 heterozygous mutants could not be analyzed later than E13.5, due to hematopoietic defects caused by Cre-induced apoptosis [36] and it was not possible to analyze conditional Tbx5 null embryos using the inducible CreER in vivo due to lethal heart defects. To circumvent these limitations, we used the Tbx4cre allele, which is expressed in the lung and trachea but not in the heart [37]. From this allele, Cre is expressed in the majority of cells of the developing lung and trachea mesenchyme, as seen with lacZ reporter expression at E13.5 (Figure 3I, 3J) [38].
Lung-specific Tbx5 null mutants, carrying a single copy of Tbx4cre, showed a range of phenotypes from an apparently normal lung to a severe decrease in lung size (data not shown). We hypothesize that the variability in phenotype is due to variable extent of recombination of the Tbx5fl allele (Figure S1). All lung-specific Tbx4 heterozygous;Tbx5 null pups (n = 13/13 from 5 litters) became cyanotic at birth and died shortly thereafter due to respiratory distress. Unlike the variable lung size in the lung-specific Tbx5 null mutants, the lungs of these mutants were consistently smaller than controls at E13.5 (Figure 3K, 3L) and at birth (Figure 3N, 3O) and had significantly fewer branching tips at E13.5 (Figure 3M). At P0 histology of the lung-specific Tbx4 heterozygous;Tbx5 null lungs is comparable to controls although the mutant tracheas show accumulation of a mucus like substance (Figure 3P, 3Q). Epithelia of mutant lungs show expression of T1α and Pro-surfactant protein C (Pro-SPC) (Figure S2A–S2D), markers for alveolar cell differentiation [10], [19], suggesting that appropriate differentiation of the lung epithelium occurs in the lung-specific Tbx4 heterozygous;Tbx5 null lungs. These lungs show lobation defects very similar to the conditional Tbx4 null;Tbx5 heterozygous mutant lungs. The accessory lobe was absent in most embryos but when present showed less branching; the cranial and caudal lobes also showed decreased branching. The cranial, medial and caudal lobes were not separated (Figure S3A, S3B). In addition, although tertiary dorsal branches were present in these lungs, they were crowded together and the secondary lateral branches had outgrown a shorter distance compared to controls (Figure S3C, S3D)
Using Tbx4cre, it is not possible to study conditional double nulls as Tbx4 is also expressed from this allele. Thus, to further explore branching morphogenesis in conditions where both alleles of Tbx4 and Tbx5 could be deleted, a lung bud culture system was used in which lung buds from embryos with or without the CreER transgene were cultured in the presence of 4-OH tamoxifen. Conditional Tbx4 null;Tbx5 heterozygous, or conditional Tbx4 heterozygous;Tbx5 null lung buds showed reduced branching (Figure 4A, 4B, 4C) consistent with the reduced number of branching tips observed in vivo. The conditional double null lungs showed a complete branching arrest by 3 days of culture (Figure 4D, 4E, 4F). The existing branches continued to elongate as seen at 4 days of culture (arrow in Figure 4E). Therefore, Tbx4 and Tbx5 are essential for continuing branching morphogenesis ex vivo.

Loss of Tbx4 and Tbx5 affects the Fgf10 signaling pathway and Wnt2 expression
Expression of the lung mesenchymal marker Fgf10 as well as epithelial targets of the Fgf10 signaling pathway, Bmp4, Spry2 and Etv5 [39], [40], [41], was analyzed in lungs with reduced Tbx4 and Tbx5 expression. Fgf10 is expressed in mesenchyme surrounding the distal epithelial tips that mark the site of future bud formation (Figure 5A) [42]. Consistent with the smaller overall lung size, there were fewer foci of Fgf10 expression in the conditional Tbx4 null;Tbx5 heterozygous lungs (Figure 5B) and in the lung-specific Tbx4 heterozygous;Tbx5 nulls (Figure 5C). The primary receptor for this pathway, Fgfr2 [43], is expressed normally in the epithelium of lung-specific Tbx4 heterozygous;Tbx5 null lungs (Figure 5D, 5E). Bmp4 (Figure 5F, 5G, 5H) and Spry2 (Figure 5I, 5J) were downregulated in Tbx4 and Tbx5-deficient lungs but Etv5 (Figure 5N, 5O) expression was not affected, although it was drastically reduced in conditional double nulls cultured ex vivo (see Figure 6J, 6K).


Wnt2, which is normally expressed in the developing lung mesenchyme (Figure 5K) [5], was greatly reduced at E13.5 in conditional Tbx4 null;Tbx5 heterozygous (Figure 5L) and lung-specific Tbx4 heterozygous;Tbx5 null lungs (Figure 5M). In addition to Fgf10 and Wnt2 signaling pathways, Shh signaling has also been implicated in branching morphogenesis in the lung [11]. Epithelial Shh (Figure 5S, 5T) and its mesenchymal receptor Ptc (Figure 5U, 5V) showed normal expression in lung-specific Tbx4 heterozygous;Tbx5 null lungs. The epithelial marker Nkx2.1, which is necessary for lung branching [44] and showed unilateral expression in the conditional Tbx5 null foreguts at the time of specification (Figure 2), showed expression similar to controls at E13.5 in the lung-specific Tbx4 heterozygous;Tbx5 null lungs (Figure 5W, 5X). At E13.5, expression of the vascular marker Pecam indicated normal development of vessels around individual bronchioles of Tbx4 and Tbx5-deficient mutants (Figure 5P, 5Q, 5R).
Tbx4 and Tbx5 interact with Fgf10 during lung development
Since mesenchymal Fgf10 expression and expression of epithelial targets was affected in mutants with reduced Tbx4 and Tbx5, we investigated the interactions between Tbx4, Tbx5 and Fgf10 during branching morhphogenesis using double and triple heterozygotus mutants. Tbx4;Tbx5 double heterozygous lungs were smaller than control lungs at E18.5 (Figure 6A, 6B). Tbx4;Tbx5;Fgf10 triple heterozygous lungs were smaller than Tbx4;Tbx5 double heterozygous lungs (Figure 6B, 6C), suggesting that Tbx4 and Tbx5 genetically interact with the Fgf10 signaling pathway during lung development. Removing one copy of Fgf10 from lung-specific Tbx4 heterozygous;Tbx5 nulls did not reduce lung size further (Figure 6D, 6E).
Despite the genetic interaction, the addition of exogenous Fgf10 to lung bud cultures of conditional double nulls did not rescue branching (Figure 6F–6I). The lack of change in Etv5 expression, an Fgf10 target, indicated that the Fgf10 signaling pathway was not activated in the presence of exogenous Fgf10 in the conditional double null lungs (Figure 6J–6M). To ensure that the Fgf10 used for the rescue experiments was active, Fgf10 coated heparin beads were placed near the branching tips of lung explants. The tips of both control and conditional double null lungs swelled up in response to Fgf10 beads but not BSA-coated heparin beads (Figure 6N, 6O) [45]. Since Fgfr2 is expressed normally in the Tbx4 and Tbx5-deficient lungs (Figure 5E) and the conditional double null lungs (data not shown), it is not surprising that the conditional double null lung tips can respond to Fgf10. However the conditional double null lungs fail to undergo branching in the presence of Fgf10. Thus, in addition to Fgf10 there must be other factors under the control of Tbx4 and Tbx5 important for activation of the Fgf10 signaling pathway leading to branching morphogenesis.
Loss of Tbx4 and Tbx5 causes tracheal/bronchial ring defects
To analyze the development of the tracheal/bronchial cartilage in embryos with reduced Tbx4 and Tbx5, cartilage rings were visualized at birth using alcian blue staining. Lung-specific Tbx5 nulls and lung-specific Tbx4 heterozygous;Tbx5 null embryos had defective cartilage ring development (Figure 7A–7C) with some normal rings (arrows in Figure 7B, 7C) and isolated foci of cartilage (black arrowheads in Figure 7B, 7C). The tracheal and bronchial lumen of newborn pups was expanded in the controls (Figure 7D, 7E) but collapsed in lung-specific Tbx4 heterozygous;Tbx5 nulls, and contained a mucus-like substance (Figure 7F, 7G). The tracheal epithelium of these mutants showed an increase in the number of mucus-producing cells, as seen by alcian blue staining (Figure 7D′, 7D″, 7F′, 7F″) [19]. To assess the development of cartilage rings at earlier stages, Sox9 expression was analyzed at E12.5 and E13.5 in lung-specific Tbx4 heterozygous;Tbx5 nulls. At E12.5, these mutant tracheas have Sox9 expression on the ventral aspect of the trachea similar to controls (Figure 7H, 7I) but fail to form mesenchymal condensations at E13.5 (Figure 7J, 7K). Expression of two genes genetically downstream of Sox9, Sox6 (Figure 7L, 7M) and Sox5 (Figure 7N, 7O), was downregulated at E13.5 whereas Col2α1, a Sox9 target, was expressed at apparently normal levels in those rings that were present (Figure 7P, 7Q). The smooth muscle marker SM22α was analyzed to assess the development of the dorsal trachealis muscle. SM22α was expressed in an expanded domain and there was a loss of the characteristic banding pattern in the mutant tracheas (Figure 7R, 7S) indicating a disruption in smooth muscle formation due to loss of Tbx4 and Tbx5.

Tbx4 and Tbx5 do not interact with Fgf10 during trachea development
Tracheas of controls (Figure 8A), Tbx4;Fgf10 double heterozygotes (Figure 8B) and Tbx5;Fgf10 double heterozygotes (Figure 8C) showed a normal pattern of 10–11 cartilage rings, suggesting a lack of genetic interactions between Tbx4 or Tbx5 and Fgf10 in trachea formation. Fgf10 null mutants do not form lungs but have a truncated trachea with 6–8 cartilage rings, some of which are aberrantly formed [22] (Figure 8D). Tbx4;Tbx5 double heterozygous mice also have tracheas with 6–8 cartilage rings but in addition have main stem bronchial cartilage rings (Figure 8E). Removing a copy of Fgf10 in these double heterozygotes did not alter the phenotype (Figure 8F) showing an Fgf10-independent role for Tbx4 and Tbx5 in the formation of tracheal/bronchial cartilage rings. Also, removing a copy of Fgf10 in the lung-specific Tbx4 heterozygous;Tbx5 nulls did not alter the phenotype of the tracheal/bronchial cartilage rings (Figure 8G, 8H). Thus, Tbx4 and Tbx5 affect lung development via control of Fgf10 expression but affect tracheal/bronchial cartilage development independently of the Fgf10 signaling pathway.

Discussion
Loss of Tbx5 affects specification of the lung buds and trachea
Tbx5 is expressed around the lung/trachea primordia at the same time that Nkx2.1, a marker of specification, is first expressed in the primordia of the foregut endoderm. Tbx4 is expressed slightly later at the time of lung bud formation. In our study, loss of Tbx5, but not Tbx4, leads to unilateral loss of lung bud specification, indicating that Tbx5 has a distinct function in lung bud formation. In contrast, in the chick, although ectopic expression of Tbx4 in the esophagus can specify lung fate, expression of a dominant negative form of Tbx4 leads to a lack of primary budding in only a third of the mutants analyzed [30]. However, the dominant negative Tbx4 could also be affecting the expression of Tbx5 targets as the repressor construct utilized the complete T-box domain and Tbx4 and Tbx5 have 94% amino acid identity in their T-box domains [46]. This interpretation is compatible with our hypothesis that Tbx5 plays a distinct role in vertebrate lung primordia specification (Figure 9A). Further our results suggest that Tbx5 regulates specification by regulating the activity of Wnt2 and Wnt2b.

Loss of Tbx5 alone leads to a lack of tracheal specification ex vivo, a phenotype similar to that of mice mutant for Bmpr1 and Bmpr2. In these mutants, although lung bud specification occurs, the ventral foregut fails to acquire tracheal identity [47]. Therefore, Tbx5 either acts in parallel with or in the BMP signaling pathway in specification of the foregut into a trachea (Figure 9A).
Tbx4 and Tbx5 interactions in lung development
In lung branching morphogenesis, Tbx4 and Tbx5 genetically interact with one another (Figure 9B). Although, neither Tbx5 (data not shown) nor Tbx4 single heterozygous lungs show a branching defect, double heterozygous lungs are smaller at E13.5 and E18.5 with a reduced number of branching tips at E13.5. Because these genes are closely related [46], they could potentially regulate the same target genes by binding to a similar T-box binding element, independently of one another. Alternatively, Tbx4 and Tbx5 could physically interact with each other as heterodimers to activate or repress transcription of downstream targets, as has been suggested for other T-box genes [48]. Even though Tbx4 and Tbx5 have a shared role in regulating lung branching, their relative contributions may not be equal, as Tbx4;Tbx5 double heterozygous lungs are smaller than conditional Tbx4 nulls. Unequal and distinct functions could be explained by domains outside of the T-box. For instance, the C terminal domain of both Tbx4 and Tbx5 has transcription activating capability which has been correlated to the shared limb outgrowth promoting activity of these genes [49] but Tbx4 also has a C terminal repressor domain which is proposed to be responsible for its distinct hind limb specific patterning activity [50]. In another example Tbx5 and Tbx4 both bind to a distinct LIM domain repeat of the LMP4 protein in the chick [51], illustrating distinct protein-protein interactions which could have functional implications.
In conditional Tbx4 nulls;Tbx5 heterozygous and lung-specific Tbx4 heterozygous;Tbx5 null lungs, reduction of Tbx4 or Tbx5 leads to a severe decrease in branching, a defect in formation of the lobes and failure of lobe separation, while absence of Tbx4 and Tbx5 leads to branching arrest. These results are in line with the published observations that antisense RNAs used against both Tbx4 and Tbx5 inhibit lung branching in culture and that this affect is explained by loss of Fgf10 expression [29]. Tbx5 binds and activates the Fgf10 promoter in vitro suggesting that Fgf10 is a direct downstream target [52]. The drastic effect on lung branching in our study is explained by loss of expression of the important regulatory genes Fgf10 and Wnt2 in the lung mesenchyme. The Fgf10 targets Bmp4 and Spry2 show very low levels of expression in the epithelium of Tbx4 and Tbx5-deficient mutants and Etv5, another Fgf10 target, shows greatly reduced expression in the conditional double null lungs, leading to the hypothesis that the Fgf10 signaling pathway is activated downstream of Tbx4 and Tbx5 in the developing lung and that Fgf10 genetically interacts with Tbx4 and Tbx5. Lungs that are triple heterozygous for Tbx4, Tbx5 and Fgf10 are reduced in size compared to Tbx4;Tbx5 double heterozygous lungs, which supports this hypothesis. Lung-specific Tbx4 heterozygote;Tbx5 null lungs are severely retarded but lung size is not affected by further loss of Fgf10, demonstrating epistasis and supporting the hypothesis that Fgf10 lies downstream of Tbx4 and Tbx5. Lack of rescue of branch formation and lack of activation of the Fgf10 signaling pathway by exogenously supplied Fgf10 in culture suggests that there are other factors downstream of Tbx4 and Tbx5 that affect Fgf10 signaling. Although, lungs deficient in Tbx4 and Tbx5 retain the ability to respond to Fgf10 coated beads (our study and [29]), they fail to activate additional factors necessary for branching morphogenesis (X in Figure 9B).
One possibility for such a factor is a mesenchymal signaling molecule that communicates with the epithelium and activates downstream target(s) required for the activation of the Fgf10 pathway. We examined expression of Ptc and Shh to determine whether the Shh pathway was involved, but Shh signaling is not affected in the lung-specific Tbx4 heterozygous;Tbx5 nulls or in the conditional double null cultures (data not shown). The extracellular matrix (ECM) molecules, heparan sulfate (HS) proteoglycans aid in Fgf10-Fgfr2 interactions during lung development [45] and hence are good candidates for factors missing in the Tbx4 and Tbx5-deficient mutants. Inhibition of heparanase decreases submandibular gland morphogenesis in culture due to deficiency of Fgf signaling [53]. Additionally, HS-deficient null mouse embryos fail to respond to Fgf signaling and the spatiotemporal expression of cell surface-tethered HS chains regulates the local reception of Fgf-signaling activity during embryonic development [54]. Tbx4 regulates ECM molecules in the developing allantois, specifically the chondroitin sulfate proteoglycan versican (R. Arora and V. E. Papaioannou unpublished observations), suggesting that ECM might be one of the targets for T-box genes in regulating development of other organs as well.
Tbx4 and Tbx5 interactions in tracheal/bronchial cartilage development
Appropriate dorsal smooth muscle development and ventral tracheal/bronchial cartilage development is important for the normal functioning of the trachea. Smooth muscle provides tracheal flexibility and the cartilaginous rings prevent tracheal collapse. Defects in trachea formation may result in tracheomalacia or tracheal stenosis. Reduction of Tbx4 and Tbx5 causes defects in both formation of the tracheal cartilage rings and development of the trachealis smooth muscle indicating interactions between Tbx4 and Tbx5. Tbx4 and Tbx5 heterozygous tracheas have 10–11 cartilage rings but Tbx4;Tbx5 double heterozygous tracheas have 6–8 cartilage rings, some of which are incomplete. Additional reduction of Tbx4 and Tbx5 in the lung-specific Tbx4 heterozygous;Tbx5 null lungs leads to a complete disruption of cartilage ring formation supporting a genetic interaction between Tbx4 and Tbx5 in trachea formation.
Sox9, a master regulator of the process of chondrogenesis, is expressed normally at E12.5 throughout the ventral mesenchyme in lung-specific Tbx4 heterozygous;Tbx5 null tracheas but at E13.5 these tracheas show a lack of characteristic mesenchymal condensations and reduced Sox9 expression. Either Tbx4 and Tbx5 control of Sox9 expression becomes more sensitive to dosage at E13.5 or Tbx4 and Tbx5 control another factor, possibly an ECM molecule, which is important for formation of mesenchymal condensations and, in the absence of these condensations, there is a down regulation of Sox9 expression (Figure 9C). Expression of Sox5 and Sox6, genes genetically downstream of Sox9 and important for condensation and cartilage formation, is downregulated in lung-specific Tbx4 heterozygous;Tbx5 null tracheas, concordant with an aberration in the process of chondrogenesis.
While Tbx4 and Tbx5 regulate lung branching by controlling Fgf10 signaling, their control of tracheal/bronchial cartilage formation is independent of Fgf10 signaling. Fgf10 homozygous mutants have 6–8 tracheal cartilage rings, although the spacing between them is reduced and the rings do not always form the characteristic C shape [8], [9], [22]. Neither Tbx4;Fgf10 double heterozygous tracheas nor Tbx5;Fgf10 double heterozygous tracheas show cartilage condensation defects or a reduction in the number of cartilage rings. In contrast, Tbx4;Tbx5 double heterozygotes show a shorter trachea with 6–8 tracheal cartilage rings suggesting Tbx4 and Tbx5 interact with each other during trachea formation but do not interact with Fgf10. The triple heterozygous tracheas do not show an exacerbation of the Tbx4;Tbx5 double heterozygous trachea phenotype. Additionally, aberrant Fgf10 signaling has been shown to affect tracheal cartilage formation and loss of Fgf10 affects Shh expression but not Sox9 expression [22]. In contrast, in the Tbx4 and Tbx5-deficient mutants Shh expression appears to be unaffected whereas Sox9 expression is reduced at E13.5. Hence, Tbx4 and Tbx5 control tracheal/bronchial cartilage formation via Sox9, independent of the Fgf10 signaling pathway.
Materials and Methods
Mouse strains, crosses, and embryo collection
Mice carrying the following alleles were genotyped as previously described: a Tbx4 conditional ‘floxed’ allele, Tbx4tm1.2Pa [55], hereafter referred to as Tbx4fl; a Tbx5 conditional floxed allele, Tbx5tm1.2Jse [56], hereafter referred to as Tbx5fl; an Fgf10 null allele [8]; ROSA26CRE-ERT2, a ubiquitous tamoxifen-inducible cre transgene [57], hereafter referred to as CreER; Tbx4-cre, an insertion into the endogenous Tbx4 allele resulting in a bicistronic allele that expresses both cre and Tbx4 in all areas of Tbx4 expression including lung and trachea [37], [38], hereafter referred to as Tbx4cre; and a R26RlacZ reporter [58]. All lines of mice were kept on mixed genetic backgrounds. Embryos were dissected from timed matings and yolk sacs were removed for PCR genotyping. The dark period was 19.00 to 05.00 h and noon on the day of a mating plug was identified as E0.5. All mouse work was carried out under Columbia University Medical Center Institutional Animal Care and Use Committee guidelines.
Tamoxifen injections
Tamoxifen (Sigma) at a concentration of 20 mg/ml in sunflower oil (Sigma) was administered to pregnant females by intraperitoneal injection between 15.30 and 19.30 hours on E8.5 or between 23.00 and 24.00 hours on E9.0.
In situ hybridization, immunohistochemistry, and histology
Whole-mount ISH, immunohistochemistry (IHC), immunofluorescence (IF) and ISH on cryosections was performed as described previously [59], [60], [61]. Primary antibodies used were anti-PECAM (Pharmingen, catalog number 01951D), anti-E-cadherin (Takara clone ECCD-2), anti T1α (Developmental Studies Hybridoma Bank antibody 8.1.1) and anti Prosurfactant Protein C (Millipore catalog number AB3786). All secondary antibodies were either peroxidase-conjugated donkey IgG from Jackson Immunochemicals or Alexa Fluor 488 from Invitrogen.
For histology embryos were removed from the uterine horns, dissected out of the decidua and fixed in Bouin's fixative (Sigma). After dehydration in ethanol, embryos were embedded in paraffin wax, sectioned at 8 µm thickness and stained with hematoxylin and eosin (H & E).
Alcian blue staining
Alcian blue staining was performed according to standard protocols [62]. Lungs and trachea were dissected out at different stages, fixed in Bouin's fixative and washed with 70% ethanol. The tissue was then equilibrated in 5% acetic acid and stained with 0.05% alcian blue in 5% acetic acid for 2 hours. The tissue was washed in 5% acetic acid to remove excess stain and dehydrated in 100% methanol, cleared in BABB (benzyl alcohol, benzyl benzoate) and photographed. For alcian blue staining on sections, 8 µm paraffin sections were rehydrated, treated with 0.05% alcian blue in 5% acetic acid and then counterstained with nuclear fast red.
Foregut and lung bud culture
Foregut culture was carried out as described previously [34]. Foreguts were isolated from 8–16 somite stage embryos using tungsten needles and cultured at 37°C in the presence of 95% air and 5% CO2 on Transwell-Col filters (Fisher Scientific) containing 1.5 ml BGJb media (Invitrogen) supplemented with 10% fetal bovine serum (FBS), 0.2 mg/ml vitamin C (Sigma) and 2 µM 4-OH tamoxifen (Sigma). For lung bud culture, lung buds were dissected at E11.5 in phosphate buffered saline (PBS) with 0.1% bovine serum albumin (Sigma) and cultured in media containing DMEM (Invitrogen) with 10% fetal bovine serum, 1% penicillin/streptomycin (Invitrogen) and 1 µM 4-OH tamoxifen on 3.0 µm filters (Millipore) or 0.4 µm Transwell filters (Fisher Scientific). Similar results were obtained using both types of filter; results reported are for experiments with Millipore filters. Where specified, Fgf10 (R&D) was added after 1 day of culture at a concentration of 500 ng/ml. In some experiments heparin beads coated with Fgf10 (100 µg/ml) or BSA (100 µg/ml) were placed near the branching tips of the explants after 1 day of culture. Transwell filters were used for the bead experiments.
Lung branching analysis
Lungs were stained with either E-cadherin antibody using IHC or E-cadherin RNA probe using ISH. In case of whole mount lungs, lobes were separated and photographed to count the number of branching tips. The cultured lungs were photographed and the branching tips were counted. For some experiments, after E-cadherin ISH, lungs were post fixed in 4% PFA, washed with PBT, dehydrated in 100% methanol, cleared in BABB and then photographed.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. WhitsettJAWertSETrapnellBC 2004 Genetic disorders influencing lung formation and function at birth. Hum Mol Genet 13 Spec No 2 R207 215
2. CardosoWVLuJ 2006 Regulation of early lung morphogenesis: questions, facts and controversies. Development 133 1611 1624
3. SpoonerBSWessellsNK 1970 Mammalian lung development: interactions in primordium formation and bronchial morphogenesis. J Exp Zool 175 445 454
4. MetzgerRJKleinODMartinGRKrasnowMA 2008 The branching programme of mouse lung development. Nature 453 745 750
5. GossAMTianYTsukiyamaTCohenEDZhouD 2009 Wnt2/2b and [beta]-catenin signaling are necessary and sufficient to specify lung progenitors in the foregut. Developmental Cell 17 290 298
6. Harris-JohnsonKSDomyanETVezinaCMSunX 2009 beta-catenin promotes respiratory progenitor identity in mouse foregut. Proc Natl Acad Sci U S A 106 16287 16292
7. DomyanETSunX 2011 Patterning and plasticity in development of the respiratory lineage. Dev Dyn 240 477 485
8. MinHDanilenkoDMScullySABolonBRingBD 1998 Fgf-10 is required for both limb and lung development and exhibits striking functional similarity to Drosophila branchless. Genes Dev 12 3156 3161
9. SekineKOhuchiHFujiwaraMYamasakiMYoshizawaT 1999 Fgf10 is essential for limb and lung formation. Nat Genet 21 138 141
10. WeaverMYinglingJMDunnNRBellusciSHoganBL 1999 Bmp signaling regulates proximal-distal differentiation of endoderm in mouse lung development. Development 126 4005 4015
11. PepicelliCVLewisPMMcMahonAP 1998 Sonic hedgehog regulates branching morphogenesis in the mammalian lung. Curr Biol 8 1083 1086
12. MillerLAWertSEClarkJCXuYPerlAK 2004 Role of Sonic hedgehog in patterning of tracheal-bronchial cartilage and the peripheral lung. Dev Dyn 231 57 71
13. MendelsohnCLohnesDDecimoDLufkinTLeMeurM 1994 Function of the retinoic acid receptors (RARs) during development (II) Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 120 2749 2771
14. DeLiseAMFischerLTuanRS 2000 Cellular interactions and signaling in cartilage development. Osteoarthritis Cartilage 8 309 334
15. de CrombruggheBLefebvreVNakashimaK 2001 Regulatory mechanisms in the pathways of cartilage and bone formation. Curr Opin Cell Biol 13 721 727
16. AkiyamaHChaboissierMCMartinJFSchedlAde CrombruggheB 2002 The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes Dev 16 2813 2828
17. HardinghamTEOldershawRATewSR 2006 Cartilage, SOX9 and Notch signals in chondrogenesis. J Anat 209 469 480
18. ParkJZhangJJMoroAKushidaMWegnerM 2010 Regulation of Sox9 by Sonic Hedgehog (Shh) is essential for patterning and formation of tracheal cartilage. Dev Dyn 239 514 526
19. QueJLuoXSchwartzRJHoganBL 2009 Multiple roles for Sox2 in the developing and adult mouse trachea. Development 136 1899 1907
20. VermotJNiederreitherKGarnierJMChambonPDolleP 2003 Decreased embryonic retinoic acid synthesis results in a DiGeorge syndrome phenotype in newborn mice. Proc Natl Acad Sci U S A 100 1763 1768
21. TiozzoCDe LangheSCarraroGAlamDANagyA 2009 Fibroblast growth factor 10 plays a causative role in the tracheal cartilage defects in a mouse model of Apert syndrome. Pediatr Res 66 386 390
22. SalaFGDel MoralPMTiozzoCAlamDAWarburtonD 2011 FGF10 controls the patterning of the tracheal cartilage rings via Shh. Development 138 273 282
23. NaicheLAHarrelsonZKellyRGPapaioannouVE 2005 T-box genes in vertebrate development. Annu Rev Genet 39 219 239
24. Gibson-BrownJJAgulnikSISilverLMPapaioannouVE 1998 Expression of T-box genes Tbx2-Tbx5 during chick organogenesis. Mech Dev 74 165 169
25. ChapmanDLGarveyNHancockSAlexiouMAgulnikSI 1996 Expression of the T-box family genes, Tbx1-Tbx5, during early mouse development. Dev Dyn 206 379 390
26. JeromeLAPapaioannouVE 2001 DiGeorge syndrome phenotype in mice mutant for the T-box gene, Tbx1. Nat Genet 27 286 291
27. NaicheLAPapaioannouVE 2003 Loss of Tbx4 blocks hindlimb development and affects vascularization and fusion of the allantois. Development 130 2681 2693
28. BruneauBGNemerGSchmittJPCharronFRobitailleL 2001 A murine model of Holt-Oram syndrome defines roles of the T-box transcription factor Tbx5 in cardiogenesis and disease. Cell 106 709 721
29. Cebra-ThomasJABromerJGardnerRLamGKSheipeH 2003 T-box gene products are required for mesenchymal induction of epithelial branching in the embryonic mouse lung. Dev Dyn 226 82 90
30. SakiyamaJYamagishiAKuroiwaA 2003 Tbx4-Fgf10 system controls lung bud formation during chicken embryonic development. Development 130 1225 1234
31. TsengYRSuYNLuFLJengSFHsiehWS 2007 Holt-Oram syndrome with right lung agenesis caused by a de novo mutation in the TBX5 gene. Am J Med Genet A 143A 1012 1014
32. ElluruRGWhitsettJA 2004 Potential role of Sox9 in patterning tracheal cartilage ring formation in an embryonic mouse model. Arch Otolaryngol Head Neck Surg 130 732 736
33. BadriKRZhouYSchugerL 2008 Embryological origin of airway smooth muscle. Proc Am Thorac Soc 5 4 10
34. ChenFCaoYQianJShaoFNiederreitherK 2010 A retinoic acid-dependent network in the foregut controls formation of the mouse lung primordium. J Clin Invest 120 2040 2048
35. DesaiTJMalpelSFlentkeGRSmithSMCardosoWV 2004 Retinoic acid selectively regulates Fgf10 expression and maintains cell identity in the prospective lung field of the developing foregut. Dev Biol 273 402 415
36. NaicheLAPapaioannouVE 2007 Cre activity causes widespread apoptosis and lethal anemia during embryonic development. Genesis 45 768 775
37. LuriaVKrawchukDJessellTMLauferEKaniaA 2008 Specification of motor axon trajectory by ephrin-B:EphB signaling: symmetrical control of axonal patterning in the developing limb. Neuron 60 1039 1053
38. NaicheLAAroraRKaniaALewandoskiMPapaioannouVE 2011 Identity and fate of Tbx4-expressing cells reveal developmental cell fate decisions in the allantois, limb, and external genitalia. Dev Dyn 240 2290 2300
39. WeaverMDunnNRHoganBL 2000 Bmp4 and Fgf10 play opposing roles during lung bud morphogenesis. Development 127 2695 2704
40. HorowitzASimonsM 2008 Branching morphogenesis. Circ Res 103 784 795
41. FirnbergNNeubuserA 2002 FGF signaling regulates expression of Tbx2, Erm, Pea3, and Pax3 in the early nasal region. Dev Biol 247 237 250
42. BellusciSGrindleyJEmotoHItohNHoganBL 1997 Fibroblast growth factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 124 4867 4878
43. ArmanEHaffner-KrauszRGorivodskyMLonaiP 1999 Fgfr2 is required for limb outgrowth and lung-branching morphogenesis. Proc Natl Acad Sci U S A 96 11895 11899
44. KimuraSHaraYPineauTFernandez-SalgueroPFoxCH 1996 The T/ebp null mouse: thyroid-specific enhancer-binding protein is essential for the organogenesis of the thyroid, lung, ventral forebrain, and pituitary. Genes Dev 10 60 69
45. IzvolskyKIShoykhetDYangYYuQNugentMA 2003 Heparan sulfate-FGF10 interactions during lung morphogenesis. Dev Biol 258 185 200
46. PapaioannouVEGoldinSN 2008 Introduction to the T-box genes and their roles in develoopmental signaling pathways. EpsteinCJEricksonRPWynshaw-BorisA Inborn Errors of Development. The Molecular Basis of Clinical Disorders of Morphogenesis, 2nd Edition Oxford Oxford University Press 852 861
47. DomyanETFerrettiEThrockmortonKMishinaYNicolisSK 2011 Signaling through BMP receptors promotes respiratory identity in the foregut via repression of Sox2. Development 138 971 981
48. GoeringLMHoshijimaKHugBBisgroveBKispertA 2003 An interacting network of T-box genes directs gene expression and fate in the zebrafish mesoderm. Proc Natl Acad Sci U S A 100 9410 9415
49. DubocVLoganMP 2011 Regulation of limb bud initiation and limb-type morphology. Dev Dyn 240 1017 1027
50. OuimetteJFJolinMLL'HonoreAGifuniADrouinJ 2010 Divergent transcriptional activities determine limb identity. Nat Commun 1 35
51. KrauseAZachariasWCamarataTLinkhartBLawE 2004 Tbx5 and Tbx4 transcription factors interact with a new chicken PDZ-LIM protein in limb and heart development. Dev Biol 273 106 120
52. AgarwalPWylieJNGalceranJArkhitkoOLiC 2003 Tbx5 is essential for forelimb bud initiation following patterning of the limb field in the mouse embryo. Development 130 623 633
53. PatelVNKnoxSMLikarKMLathropCAHossainR 2007 Heparanase cleavage of perlecan heparan sulfate modulates FGF10 activity during ex vivo submandibular gland branching morphogenesis. Development 134 4177 4186
54. ShimokawaKKimura-YoshidaCNagaiNMukaiKMatsubaraK 2011 Cell surface heparan sulfate chains regulate local reception of FGF signaling in the mouse embryo. Dev Cell 21 257 272
55. NaicheLAPapaioannouVE 2007 Tbx4 is not required for hindlimb identity or post-bud hindlimb outgrowth. Development 134 93 103
56. MoriADZhuYVahoraINiemanBKoshiba-TakeuchiK 2006 Tbx5-dependent rheostatic control of cardiac gene expression and morphogenesis. Dev Biol 297 566 586
57. de LucaCKowalskiTJZhangYElmquistJKLeeC 2005 Complete rescue of obesity, diabetes, and infertility in db/db mice by neuron-specific LEPR-B transgenes. J Clin Invest 115 3484 3493
58. SorianoP 1999 Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat Genet 21 70 71
59. WilkinsonDGNietoMA 1993 Detection of messenger RNA by in situ hybridization to tissue sections and whole mounts. Methods Enzymol 225 361 373
60. DavisCA 1993 Whole-mount immunohistochemistry. Methods Enzymol 225 502 516
61. GrieshammerULeMPlumpASWangFTessier-LavigneM 2004 SLIT2-mediated ROBO2 signaling restricts kidney induction to a single site. Dev Cell 6 709 717
62. NagyAGertsensteinMVinterstenKBehringerR 2009 Alcian blue staining of the mouse fetal cartilaginous skeleton. Cold Spring Harbor Protocols 2009 pdb.prot5169
63. DouglasNCHengKSauerMVPapaioannouVE 2011 Dynamic expression of Tbx2 subfamily genes in development of the mouse reproductive system. Dev Dyn 241 365 375
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 8
Nejčtenější v tomto čísle
- Dissecting the Gene Network of Dietary Restriction to Identify Evolutionarily Conserved Pathways and New Functional Genes
- It's All in the Timing: Too Much E2F Is a Bad Thing
- A Quantitative Comparison of the Similarity between Genes and Geography in Worldwide Human Populations
- Variation of Contributes to Dog Breed Skull Diversity