
Deciphering the Transcriptional-Regulatory Network of Flocculation in
In the fission yeast Schizosaccharomyces pombe, the transcriptional-regulatory network that governs flocculation remains poorly understood. Here, we systematically screened an array of transcription factor deletion and overexpression strains for flocculation and performed microarray expression profiling and ChIP–chip analysis to identify the flocculin target genes. We identified five transcription factors that displayed novel roles in the activation or inhibition of flocculation (Rfl1, Adn2, Adn3, Sre2, and Yox1), in addition to the previously-known Mbx2, Cbf11, and Cbf12 regulators. Overexpression of mbx2+ and deletion of rfl1+ resulted in strong flocculation and transcriptional upregulation of gsf2+/pfl1+ and several other putative flocculin genes (pfl2+–pfl9+). Overexpression of the pfl+ genes singly was sufficient to trigger flocculation, and enhanced flocculation was observed in several combinations of double pfl+ overexpression. Among the pfl1+ genes, only loss of gsf2+ abrogated the flocculent phenotype of all the transcription factor mutants and prevented flocculation when cells were grown in inducing medium containing glycerol and ethanol as the carbon source, thereby indicating that Gsf2 is the dominant flocculin. In contrast, the mild flocculation of adn2+ or adn3+ overexpression was likely mediated by the transcriptional activation of cell wall–remodeling genes including gas2+, psu1+, and SPAC4H3.03c. We also discovered that Mbx2 and Cbf12 displayed transcriptional autoregulation, and Rfl1 repressed gsf2+ expression in an inhibitory feed-forward loop involving mbx2+. These results reveal that flocculation in S. pombe is regulated by a complex network of multiple transcription factors and target genes encoding flocculins and cell wall–remodeling enzymes. Moreover, comparisons between the flocculation transcriptional-regulatory networks of Saccharomyces cerevisiae and S. pombe indicate substantial rewiring of transcription factors and cis-regulatory sequences.
Published in the journal:
. PLoS Genet 8(12): e32767. doi:10.1371/journal.pgen.1003104
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1003104
Summary
In the fission yeast Schizosaccharomyces pombe, the transcriptional-regulatory network that governs flocculation remains poorly understood. Here, we systematically screened an array of transcription factor deletion and overexpression strains for flocculation and performed microarray expression profiling and ChIP–chip analysis to identify the flocculin target genes. We identified five transcription factors that displayed novel roles in the activation or inhibition of flocculation (Rfl1, Adn2, Adn3, Sre2, and Yox1), in addition to the previously-known Mbx2, Cbf11, and Cbf12 regulators. Overexpression of mbx2+ and deletion of rfl1+ resulted in strong flocculation and transcriptional upregulation of gsf2+/pfl1+ and several other putative flocculin genes (pfl2+–pfl9+). Overexpression of the pfl+ genes singly was sufficient to trigger flocculation, and enhanced flocculation was observed in several combinations of double pfl+ overexpression. Among the pfl1+ genes, only loss of gsf2+ abrogated the flocculent phenotype of all the transcription factor mutants and prevented flocculation when cells were grown in inducing medium containing glycerol and ethanol as the carbon source, thereby indicating that Gsf2 is the dominant flocculin. In contrast, the mild flocculation of adn2+ or adn3+ overexpression was likely mediated by the transcriptional activation of cell wall–remodeling genes including gas2+, psu1+, and SPAC4H3.03c. We also discovered that Mbx2 and Cbf12 displayed transcriptional autoregulation, and Rfl1 repressed gsf2+ expression in an inhibitory feed-forward loop involving mbx2+. These results reveal that flocculation in S. pombe is regulated by a complex network of multiple transcription factors and target genes encoding flocculins and cell wall–remodeling enzymes. Moreover, comparisons between the flocculation transcriptional-regulatory networks of Saccharomyces cerevisiae and S. pombe indicate substantial rewiring of transcription factors and cis-regulatory sequences.
Introduction
Flocculation is an inherent characteristic of yeasts involving asexual aggregation of cells into flocs that separate rapidly from the medium (reviewed recently in [1], [2]). Individual yeast cells transition into this morphological state as an adaptation to various environmental stresses by shielding the inner cells of the flocs [3]. The flocculent trait has also proven highly beneficial in industrial yeast applications by allowing efficient and cost-effective removal of cells [4]. The ability of yeast strains to flocculate is dependent on the expression of specific cell surface glycoproteins known as flocculins. Cell-to-cell adhesion occurs via binding between the flocculin and surface carbohydrates in a calcium-dependent manner [5]. The bound carbohydrates consist of various sugars including mannose, glucose, and galactose that are specific to the type of flocculin and yeast species [6]–[8]. There has been considerable interest in elucidating the genetic control of flocculation to better understand this phenomenon and generate biotechnological advances in yeast-based industries.
In Saccharomyces cerevisiae, a transcriptional-regulatory network composed of interactions between transcription factors and their flocculin gene targets is central in controlling flocculation. The primary flocculins that function in flocculation are encoded by the FLO1, FLO5, FLO9, and FLO10 genes [9]–[11]. Overexpression of the individual FLO genes is sufficient to trigger flocculation [8], [12]. However, the degree of flocculation by FLO overexpression varies from FLO1 to FLO10 exhibiting the strongest to weakest flocculation, respectively. The flocculin FLO11 also exhibits weak flocculation when overexpressed [8], but its function is mainly in cell-to-surface adhesion [13], diploid pseudohyphal growth [14], and haploid invasive growth [15]. The transcription factors required for flocculation include Flo8p and Mss11p, which primarily activate FLO1 transcription [16]. The Sacc. cerevisiae laboratory strain S288C containing a nonfunctional FLO8 gene is not able to flocculate, but flocculation is restored in this strain by the overexpression of FLO8 or MSS11 [16], [17]. In addition, Sfl1p has been shown to inhibit transcription of FLO1 in the W303-1A strain and not in S288C, likely through interactions with the Ssn6p-Tup1p global repressor and components of Mediator [18], [19].
The control of flocculation is much less known in Schizosaccharomyces pombe. The ability of the heterothallic wild-type strains 972 h− and 975 h+ to flocculate has not been observed presumably because the inducing environmental conditions have not been identified. Phenotypic analysis of constitutive flocculent mutant strains show that flocculation is dependent on the presence of calcium, but unlike Sacc. cerevisiae, the flocculin-carbohydrate interactions involve galactose rather than mannose and glucose residues [7]. Moreover, the transcriptional-regulatory network governing flocculation in S. pombe remains poorly characterized. Only a single interaction between the Mbx2 MADS box transcription factor and the gsf2+ flocculin gene is currently known [20], [21]. The gsf2+ gene was initially identified as highly upregulated in response to heterologous expression of FLO8 [20]. Overexpression of gsf2+ is sufficient to trigger flocculation while its deletion abrogates the flocculent phenotype of tup12Δ, lkh1Δ, and gsf1 mutants. In addition, gsf2+ displays additional roles in cell-to-surface adhesion and invasive growth [20]. The induction of gsf2+ during flocculation and invasive growth is mediated by Mbx2 [21]. Two other transcription factors implicated in flocculation have been reported. The CSL transcription factors Cbf11 and Cbf12 play opposing roles in flocculation where mutant strains lacking cbf11+ or overexpressing cbf12+ flocculate [22]. The direct targets of these transcription factors functioning in flocculation have not been identified, but could be several putative flocculin genes that show protein sequence homology to other yeast-related proteins [23]. Indeed, these putative flocculin genes, as well as gsf2+ are transcriptionally upregulated in certain Mediator mutants that flocculate indicating that these genes are likely repressed by Mediator [24]. Similar to Sacc. cerevisiae, the global transcriptional regulators Tup11 and Tup12 function in flocculation but their influence on the expression of these flocculin genes has not been addressed [25]. Importantly, it has not been directly demonstrated that these putative flocculin genes in S. pombe actually play a role in flocculation and the identity of the transcription factors that regulate them remains unknown.
In this study, we have initiated an extensive characterization of the transcriptional-regulatory network of S. pombe flocculation by identifying the relevant transcription factors and their flocculin gene targets. Importantly, we have also determined that heterothallic wild-type S. pombe is able to flocculate when grown in rich medium containing ethanol and glycerol as a carbon source. A screen of transcription factor deletion and overexpression strains for flocculent phenotypes revealed five novel transcriptional regulators of flocculation (Rfl1, Adn2, Adn3, Sre2, Yox1) in addition to our independent finding of Mbx2, Cbf11, and Cbf12. The strongest flocculation was observed upon overexpression of mbx2+ and deletion of rfl1+ (SPBC15D4.02) which encodes an uncharacterized fungal Zn(2)-Cys(6) transcription factor. Microarray expression profiling of the mbx2OE and rfl1Δ strains revealed good overlap in the upregulation of several flocculin genes, while ChIP-chip analysis of HA-tagged Mbx2 and Rfl1 under control of the nmt41 promoter indicated that these transcription factors bound to some of the flocculin gene promoters. Nine flocculin gene targets (pfl1+–pfl9+) including gsf2+/pfl1+ were identified. The single overexpression of these genes triggered flocculation to varying degrees and cumulative effects on flocculation were observed in double overexpression experiments. Only loss of gsf2+ could abrogate the flocculent phenotype of all the transcription factor mutants indicating that gsf2+ encodes the dominant flocculin in S. pombe. Interestingly, we discovered that certain cell wall-remodeling enzymes can also function in flocculation, and some of these genes are likely regulated by the LisH transcription factors Adn2 and Adn3. In addition to the identification of target genes within the transcriptional-regulatory network, autoregulatory and inhibitory feed-forward loops involving several transcription factors were also detected. These results provide a significant insight into the transcriptional control of flocculation in S. pombe.
Results
Screening for novel transcription factors functioning in fission yeast flocculation
Our understanding of the transcriptional-regulatory network that governs flocculation in S. pombe remains limited. To further decipher this network, we sought to systematically identify transcription factors that play a role in flocculation. A list of 101 genes encoding sequence–specific transcription factors containing a bona-fide DNA-binding domain was assembled from [26] and GeneDB [27]. From this gene list, we constructed 101 nmt1-driven overexpression strains and 92 nonessential deletions in which the entire ORF was replaced with the KanMX6/NatMX6 cassette. A detailed description of the construction and phenotypic characterization of this transcription factor mutant collection will be described elsewhere (unpublished data). The transcription factor array of overexpression and deletion strains were screened for flocculation in EMM lacking thiamine and YES media, respectively. We recovered a total of eight transcription factors in which four overexpression strains (mbx2OE, adn2OE, adn3OE and cbf12OE) and four deletions (rfl1Δ, sre2Δ, yox1Δ and cbf11Δ) exhibited flocculation. These transcription factors represent positive and negative regulators of flocculation, respectively. Among these transcription factors, only the overexpression of cbf12+ and mbx2+ and deletion of cbf11+ have been reported to cause flocculation [20], [22].
The strongest flocculation was observed in the mbx2OE and rfl1Δ strains. The flocs of the rfl1Δ strain in YES medium were larger and sedimented faster than the flocs produced in the mbx2OE strain after 48 hour induction (Figure 1A). The mbx2+ gene encodes a MADS-box transcription factor which was originally isolated in a screen for genes functioning in the biosynthesis of cell surface pyruvated galactose residues [28]. Recently, Mbx2 has been shown to function in flocculation and invasive growth by regulating the flocculin gene gsf2+ [20], [21]. The rfl1+ (repressor of flocculation) gene encodes an uncharacterized fungal Zn(2)-Cys(6) transcription factor.

The flocculation exhibited by these overexpression and deletion transcription factor mutants recovered from our screens could be abolished with the addition of galactose, but not mannose or glucose (data not shown). The amount of galactose required to completely deflocculate cells depended on the degree of flocculation. For example, mbx2OE strain could be deflocculated with 2% galactose while rfl1Δ strain required 5–10 times more galactose to completely deflocculate. Reflocculation of these strains was achieved in CaCl2 or in YES medium (data not shown).
The growth conditions that trigger flocculation in heterothallic wild-type S. pombe are not well known. To identify the inducing conditions, 972 h− and 975 h+ cells were tested on different carbon sources at different cell densities for flocculation. We determined that heterothallic wild-type cells were able to flocculate when cultured for five days at an initial concentration of 1×106 cells/ml in medium containing 1% yeast extract, 3% glycerol and, 4% ethanol (referred to as flocculation-inducing medium, Figure 1B). The degree of flocculation was slightly enhanced in strains auxotrophic for leucine, uracil, and/or adenine indicating that nutrient status may also play a role in triggering flocculation (data not shown). However, these wild-type strains flocculated significantly less in flocculation-inducing medium than the mbx2OE and rfl1Δ mutants in EMM and YES media, respectively. The weaker flocculation in these strains was more easily observed in petri-dishes incubated on an orbital rotator than in test tubes. In contrast to wild type, deletion of mbx2+ did not produce any visible flocs in the flocculation-inducing medium (Figure 1B).
Fungal genes that function in flocculation are usually associated with filamentous invasive growth [17], [20]. We hypothesized that the rfl1Δ strain would exhibit hyperfilamentous invasive growth because of its strong flocculent phenotype. Indeed, the amount of cells resistant to removal from the agar by washing in the invasive assay on LNB medium with an underlayer of YE+ALU was much greater in the rfl1Δ strain than in wild type (Figure 1C). Under the microscope, the filamentous growth like those detected by Dodgson et al. [29] was observed below the agar surface for both wild type and rfl1Δ strain with the latter showing much larger and more frequent formation of filamentous growth (data not shown). Similarly, adn2+ and adn3+ which were previously observed to have defects in invasive growth when deleted were recovered in our screens as flocculent when overexpressed [29].
Mbx2 and Rfl1 are opposing transcription factors that regulate putative flocculin genes
The strongest flocculation observed in the mbx2OE and rfl1Δ strains indicated that these two genes encode the major regulators of flocculation. Therefore, we initially focused on the characterization of these two transcription factors and proceeded to identify their target genes involved in flocculation. The nmt41-driven mbx2-HA strain was subjected to microarray expression profiling with a custom-designed S. pombe 8×15 K Agilent expression microarray (Table S2). The intermediate strength nmt41 promoter was sufficient for mbx2OE flocculation and was utilized in the microarray experiments in order to reduce possible secondary transcriptional effects compared to the strong nmt1 promoter. To better distinguish the direct target genes, ChIP-chip was also carried out concurrently on the same strain using the S. pombe 4×44 K Agilent Genome ChIP-on-chip microarray (Table S3). For the rfl1+ expression profiling and ChIP-chip experiments, the flocculent deletion mutant and nmt41-driven rfl1-HA strain were used, respectively (Tables S4 and S5). The highly-induced putative target genes identified by microarray expression profiling of these transcription factor mutant strains were validated by qPCR (Table S13).
The list of genes that were induced at least two fold in the mbx2OE or rfl1Δ strain was subjected to gene ontology analysis using the Princeton GO Term Finder (http://go.princeton.edu/cgi-bin/GOTermFinder). These induced genes were highly enriched in cell wall components with p-values of 9.0e-9 and 6.3e-6 for the mbx2OE and rfl1Δ strains, respectively. Strikingly, the most-induced genes in the mbx2OE strain encoded cell surface glycoproteins. The cell surface glycoprotein genes up-regulated above two-fold were SPAC186.01, gsf2+, SPAC977.07c/SPBC1348.08c, SPCC188.09c, fta5+, SPBC947.04, SPBC359.04c, SPBC1289.15, SPAPB2C8.01, SPAC1F8.02c, SPAPB18E9.04c, SPCC553.10, and SPBPJ4664.02, which all but gsf2+ and the last 4 genes were predicted to be pombe adhesins based on BLAST sequence analysis (Figure 2A; [23]). SPAC977.07c and SPBC1348.08c are gene duplications with 100% sequence identity. To our knowledge, these genes with the exception of gsf2+ have not been characterized further. The induction of these genes in the mbx2OE strain ranged from 2 to 112-fold relative to the empty vector control (Figure 2A, Table S13). In addition, several genes (agn2+, psu1+, SPAC4H3.03c and gas2+) encoding cell wall-remodeling enzymes such as glucan glucosidases and a betaglucanosyltransferase were induced up to 91-fold compared to the empty vector control when mbx2+ was overexpressed (Figure 2A). In the rfl1Δ expression data, a similar set of cell surface glycoprotein genes were upregulated at a comparable level as the mbx2OE expression data except for SPAC1F8.02, SPBC359.04c, SPAPB18E9.04c and SPBPJ4664.02 (Figure 2A, Table S13). In contrast to the mbx2OE strain, the same genes encoding the cell wall-remodeling enzymes were not highly upregulated in the rfl1Δ strain (Figure 2A).

Of the thirteen highly-induced cell surface glycoprotein genes in the mbx2OE expression data, nine of them were detected with ChIP-chip indicating that these genes are very likely the direct transcriptional targets of Mbx2 (Figure 2A). Four of the nine highly-induced cell surface glycoprotein genes in the rfl1Δ strain were detected with ChIP-chip confirming that these genes are probably direct transcriptional targets of Rfl1 (Figure 2A). For both Mbx2 and Rfl1, gsf2+, fta5+ and SPAPB2C8.01 were detected in the expression microarray and ChIP-chip experiments (Figure 2A).
Next, we sought further evidence that these cell surface glycoprotein genes were targets of Mbx2 and Rfl1 by epistasis studies. We decided to study a subset of these genes, which included the majority of the gene sequences analyzed by Linder and Gustafsson [23], [24]. The mbx2+ gene was overexpressed in single deletions of these putative target genes and their degree of flocculation was determined visually in petri-dishes, as well as quantitatively (Table S14). The putative glycoprotein gene SPAPB15E9.01c was included in these studies, because even though the transcript was downregulated in both mbx2OE and rfl1Δ strains, ChIP-chip analysis detected Mbx2 and Rfl1 association with its promoter (Figure 2A). Deletion of gsf2+ decreased mbx2OE flocculation to the greatest extent while the reduction of flocculation was less extensive in the other single deletion mutants (Figure 2B, Table S14). The degree of reduction in mbx2OE flocculation roughly corresponded to the pfl numbers, which were assigned based on the degree of flocculation when overexpressed (see below). Moreover, mbx2OE flocculation was completely abrogated in the gsf2Δ pfl9Δ double mutant indicating that the reduction of mbx2OE flocculation in these mutants were additive in some cases (Figure 2B). Similar experiments were performed for rfl1+ in which flocculation was assayed in the same putative target deletions in the rfl1Δ background. The flocculation exhibited in the rfl1Δ strain was completely abolished by the deletion of gsf2+, but not by the deletion of pfl9+ (Figure 2C).
To further analyze the expression microarray datasets of Mbx2 and Rfl1, the promoter regions of the differentially-expressed genes were subjected to the motif-finding algorithms RankMotif++ and MEME to identify their binding specificities [30], [31]. Mbx2 is a member of the MEF2-MADS box transcription factor family which has been shown to bind to the consensus sequence 5′-(C/T)TA(T/A)4TA(G/A)-3′ [28], [32], [33]. The Mbx2 binding specificity obtained by RankMotif++ closely resembled this known consensus sequence (Figure 2D). Similarly, RankMotif++ generated an Rfl1 binding specificity that resembled known consensus sequences of several members of the fungal Zn(2)-Cys(6) transcription factor family (Figure 2E). The binding specificity of Zn(2)-Cys(6) DNA-binding domains is composed of conserved GC-rich trinucleotides spaced by a variable sequence region differing in length among members of the transcription factor family [34]. Analyses of the Mbx2 and Rfl1 expression microarray and ChIP-chip datasets by MEME did not generate any candidate DNA motifs.
Altogether, these results demonstrate that Mbx2 and Rfl1 are transcription factors responsible for regulation of flocculation in fission yeast by activating or repressing the transcription of candidate S. pombe flocculin genes, respectively.
The putative flocculin gene targets of Mbx2 and Rfl1 are sufficient to induce flocculation when overexpressed
Besides gsf2+, the other putative target genes of Mbx2 and Rfl1 that encode for cell surface glycoproteins share some amino acid sequence homology with domains found in other fungal adhesins [23]. However, the role of these glycoprotein genes in flocculation has not been demonstrated. Overexpression studies were employed to the aforementioned set of putative flocculin target genes of Mbx2 and Rfl1 to determine whether they function directly in flocculation. Each single overexpression of these flocculin genes was able to induce flocculation to varying degrees with the strongest flocculation observed in the gsf2OE strain which produced visible flocs within one day (Figure 3A; Table S14). Weaker flocculation was observed from the overexpression of the other flocculin genes after total incubation of 2–7 days in EMM minus thiamine medium with sub-culturing into fresh medium in Day 3. The flocculation images of these overexpression strains shown in Figure 3A were captured after total of 7 days of induction. As a result of these observations, we named these genes pfl+ for Pombe Flocculins and numbered them according to their degree of flocculation when overexpressed: pfl1+/gsf2+ (referred as gsf2+ hereafter), pfl2+/SPAPB15E9.01c, pfl3+/SPBC947.04, pfl4+/SPCC188.09c, pfl5+/SPBC1289.15, pfl6+/SPAC977.07c, pfl7+/SPBC359.04c, pfl8+/fta5+ (referred as fta5+ hereafter) and pfl9+/SPAC186.01. Furthermore, we overexpressed some double combinations of the weaker flocculin genes to determine whether flocculation could be additive. Indeed, the pfl4+ pfl9+, pfl6+ pfl9+, and fta5+ pfl9+ double overexpression strains flocculated earlier and formed larger flocs than their corresponding single overexpressors, thus, demonstrating the additive effect of these flocculins (Figure 3B, Table S14). We next tested the single deletions of the pfl+ genes for their ability to flocculate in flocculation-inducing medium. No visible flocculation was observed in the gsf2Δ strain while wild type was flocculent (Figure 1B). In contrast, flocculation still occurred in the pfl2Δ–pfl9Δ strains in the inducing medium indicating that gsf2+ encodes the dominant flocculin and the other flocculin genes are dispensable for flocculation (data not shown).

These observations revealed that the contribution in flocculation by these pfl+ genes varied and certain combinations of pfl+ were additive. The strength of flocculation by the single overexpression of pfl+ genes was directly correlated with the reduction of mbx2OE flocculation in the corresponding deletion strains (Figure 2B and Figure 3A, Table S14). For example, the pfl2OE strain which produced larger flocs than the pfl3OE–pfl9OE strains exhibited a greater inhibition of mbx2OE flocculation when deleted. Similarly, the flocculation of the rfl1Δ strain was completely abrogated by the deletion of gsf2+, but not at all by the deletion of pfl9+ (Figure 2C). Consistent with the above results, the deletion of both gsf2+ and pfl9+ led to a greater abrogation of mbx2OE flocculation compared to each deletion alone (Figure 2B). In summary, we have demonstrated that these pfl+ genes encode for S. pombe flocculins and Gsf2 is the dominant flocculin.
Positive and negative autoregulation of mbx2+ and rfl1+, respectively
Interestingly, ChIP-chip analysis also detected binding of Mbx2 and Rfl1 to their own promoters, as well as Rfl1 binding to the mbx2+ promoter (Figure 2A), indicating autoregulation and mbx2+ regulation by Rfl1 within the transcriptional-regulatory network of S. pombe flocculation. Mbx2 also appeared to be associated with the rfl1+ promoter, but this interaction was marginal as it was found just above the detection threshold for ChIP-chip (Figure 2A). To investigate the autoregulation of mbx2+, the gene was C-terminal tagged with GFP at its native locus (mbx2-GFP). However, the GFP-tagged strain resulted in a hypermorphic allele that displayed constitutive flocculation and nuclear localization of Mbx2-GFP (see below). We speculated that the removal of the 3′-untranslated region of mbx2+ during the C-terminal tagging may be the cause of the hypermorphic allele. To bypass this potential problem, we created an N-terminal GFP-tagged allele (GFP-mbx2) with an intact 5′-untranslated region and approximately 1 kb of native promoter sequence. In contrast to the C-terminal tagged hypermorphic allele, the N-terminal tagged GFP-Mbx2 expression was comparable to background levels and the strain did not exhibit constitutive flocculation (Figure 4A). Moreover, the GFP-mbx2 strain flocculated when grown in glycerol-inducing medium indicating that the tagged protein is functional (Table S14). When nmt1-driven mbx2+ expression was induced for 9 hours in the GFP-mbx2 strain, nuclear GFP-Mbx2 expression was detected, indicating that Mbx2 can activate its own expression (Figure 4A). As expected, this strain was now flocculent. Longer induction of nmt1-driven mbx2+ expression resulted in greater GFP-Mbx2 expression with multi-nucleated GFP foci (data not shown). The positive autoregulation of mbx2+ is likely to be direct as several putative MEF2-binding sequences (e.g. 5′-TTAAAAATAG-3′) are located within 1000 bp upstream from the mbx2+ start codon (data not shown).

To determine whether negative autoregulation occurs with rfl1+, a C-terminal GFP-tagged strain under native control was generated (rfl1-GFP). The localization of Rfl1-GFP was nuclear in the rfl1-GFP strain (Figure 4B). The induction of nmt1-driven rfl1+ expression for 18 hours in the rfl1-GFP strain led to a reduced nuclear Rfl1-GFP signal and a slightly increased cytoplasmic Rfl1-GFP signal (Figure 4B). However, overall Rfl1-GFP expression in the cell was reduced when Rfl1 was overexpressed compared to the empty vector control (Figure 4B; two-tailed t-test; p value<0.01). In contrast to our observations with the Rfl1-GFP protein expression, we found that there was no decrease of the Rfl1-GFP transcript when rfl1+ was overexpressed (Table S13). These results indicate that although Rfl1 can bind to its own promoter, negative autoregulation appears marginal or may not be occurring.
Rfl1 represses mbx2+ expression
The observation that Rfl1 is associated with the mbx2+ promoter by ChIP-chip suggests that Rfl1 may oppose Mbx2 function in flocculation by repressing its expression. To test this hypothesis, we first examined the genetic interactions between mbx2+ and rfl1+. The mbx2Δ rfl1Δ double mutant did not display flocculation indicating that mbx2+ is epistatic to rfl1+ (Figure 5A). In addition, the flocculation associated with mbx2OE was abrogated by co-overexpression of rfl1+ (Figure 5A). These results are consistent with mbx2+ being downstream of rfl1+ and that rfl1+ opposes mbx2+ function in flocculation.

We next utilized the C-terminal and N-terminal GFP-tagged mbx2+ strains to further determine if Rfl1 represses mbx2+ expression. First, Rfl1 was overexpressed in the hypermorphic C-terminal tagged mbx2-GFP allele which shows constitutive nuclear Mbx2-GFP expression and flocculation. This resulted in the near-abolishment of both the GFP signal (Figure 5B) and flocculation (data not shown) in the hypermorphic mbx2 allele. Second, when the N-terminal tagged GFP-mbx2 strain was crossed into the rfl1Δ background, the resulting strain displayed dramatic increase in nuclear GFP-Mbx2 expression (Figure 5C) and flocculation strength equivalent to the rfl1Δ strain (data not shown). These results support the hypothesis that mbx2+ expression is repressed by Rfl1 in non-flocculent cells.
Overexpression of cbf12+ causes flocculation due to up-regulation of gsf2+
Cbf12, a member of the CSL transcription factor family has previously been reported to trigger flocculation when overexpressed [22]. However, the target genes of Cbf12 that function in flocculation have not been identified. To further elucidate the role of cbf12+ in flocculation, we took a similar approach to identify its direct target genes by concurrent expression microarray profiling and ChIP-chip analysis of the nmt41-driven cbf12-HA strain (Tables S6 and S7, respectively).
When cbf12+ was deleted and cultured in flocculation-inducing medium, flocculation was abolished (Figure 6A). In contrast, overexpression of cbf12+ by the nmt1 promoter triggered flocculation (Figure 6C) and produced a bowling pin–shaped phenotype after 24 hours in medium lacking thiamine (data not shown). Further induction of the nmt1-driven cbf12+ caused the strain to become sick and granulated, eventually leading to growth arrest (data not shown). To reduce the toxic effects of cbf12+ overexpression, an nmt41-driven cbf12-HA strain was used for concurrent expression profiling and ChIP-chip analysis.

Gene ontology analysis was carried out separately on the top 50 most highly-induced genes and all 160 promoter-occupied genes by Cbf12 with the Princeton GO Term Finder. Functional enrichment of genes in cell surface (p = 1.8e-7) and plasma membrane (p = 5.7e-4) was detected for the highly-induced and promoter-occupied genes, respectively. These genes included several flocculin genes, (Figure 6B). Both gsf2+ and pfl7+ were among the five highest induced genes (18.1 and 27.6-fold, respectively) in the cbf12OE strain and were also detected by ChIP-chip (Figure 6B) suggesting that Cbf12 directly activates the transcription of gsf2+ and pfl7+ for flocculation. The flocculation triggered by cbf12+ overexpression was completely abrogated in the gsf2Δ background, whereas deletion of pfl7+ had little effect (Figure 6C, Table S14). This was consistent with the hypothesis that gsf2+ encodes the dominant flocculin. In addition, loss of gsf2+ or pfl7+ did not alter the bowling-pin cell shape or the reduced fitness phenotypes of the cbf12OE strain indicating that these two phenotypes were not due to the upregulation of the flocculin genes (data not shown). The much weaker flocculation observed in the cbf12OE strain in comparison to the mbx2OE and gsf2OE strains may be attributed to additional defects in cell and nuclear division, which would cause early growth arrest before the full flocculation potential could be reached [22].
Consistent with previous findings, C-terminal GFP-tagged Cbf12 under native control was expressed predominantly in the nucleus in stationary phase cells while expression in logarithmic cells was comparable to background (Figure 6D; [22]). Compared to logarithmic growth in rich medium, Cbf12-GFP nuclear expression increased in cells grown in flocculation-inducing medium, thus supporting its role in flocculation (Figure 6D). Interestingly, Cbf12 was also detected by ChIP-chip to bind to its own promoter (Figure 6B). Indeed, positive autoregulation appears to occur as native Cbf12-GFP expression increased greater than three-fold when nmt1-driven cbf12+ was ectopically expressed in logarithmically growing cells (Figure 6E).
Recently, it was demonstrated that an N-terminal-truncated Cbf12 bound to probes containing a canonical CSL binding motif (5′-GTGGGAA-3′) by gel mobility shift assay [35]. We next searched for a similar DNA binding sequence for Cbf12 from the expression microarray and ChIP-chip cbf12OE datasets by RankMotif++ and MEME. RankMotif++ and MEME analyses of the expression microarray and ChIP-chip data, respectively, did not identify a binding specificity for Cbf12. However, when the promoters of up-regulated genes in the cbf12OE strain belonging to the cell surface GO category were subjected to MEME analysis, a motif closely matching the canonical CSL binding motif (6/7 nucleotide match) was recovered (Figure 6F).
These results demonstrate Cbf12 as part of the transcriptional-regulatory network of fission yeast flocculation by controlling the transcription of several flocculin genes including gsf2+.
The flocculation function of yox1+, sre2+, and cbf11+ is dependent on gsf2+
From our transcription factor screens, the deletion of yox1+, sre2+, or cbf11+ also resulted in flocculation, although the size of the flocs were smaller than observed in mbx2OE, cbf12OE and rfl1Δ strains (Figure 7A, Table S14). Yox1 has been implicated in a negative autoregulatory loop to prevent inappropriate transcriptional expression of MBF gene targets, while the function of Sre2, which shows homology to the human sterol regulatory element binding protein SREBP-1A remains largely unknown [36], [37]. A role of Yox1 and Sre2 in flocculation has not been reported. In contrast, cbf11+ encodes a CSL transcription factor that plays a role in flocculation, but its target genes are not known [22].

To elucidate the transcriptional flocculation program of yox1+, sre2+ and cbf11+, expression microarray profiling was conducted on the corresponding flocculent deletion strains in rich medium (Tables S8, S9, S10). The expression microarray profiles of yox1Δ and sre2Δ most resembled each other compared to the other strains described in this study (Figure 7B). Genes upregulated by at least two-fold in the yox1Δ and sre2Δ strains showed enrichment for ribosomal subunits (p = 2.8e-31 and 7.4e-25 for yox1Δ and sre2Δ, respectively) and mitochondrial membrane transporters (p = 7.5e-5 and 1.2e-3 for yox1Δ and sre2Δ, respectively). These findings did not intuitively answer our questions as to how these two transcription factors might be related or associated with the flocculation pathway. We next examined whether any of the flocculin genes and their putative regulators were induced in the yox1Δ and sre2Δ strains. In the sre2Δ strain, gsf2+, pfl3+ and fta5+ transcripts were upregulated 3.7, 2.5 and 3.1-fold, respectively, indicating that the expression of these genes could be contributing to the flocculent phenotype (Figure 7C). In contrast, mbx2+ and cbf12+ transcripts were downregulated approximately 2-fold suggesting that the elevated levels of gsf2+, pfl3+ and fta5+ transcripts in the sre2Δ strain were not mediated by Mbx2 and Cbf12 (Figure 7C). Similarly in the yox1Δ strain, we observed that gsf2+ and pfl3+ transcripts were upregulated although less than in the sre2Δ strain, and mbx2+ and cbf12+ were also downregulated (Figure 7C). Therefore, this suggests that sre2+ and yox1+ may be involved in the repression of flocculation through a pathway independent from mbx2+ and cbf12+.
The microarray expression profile of the cbf11Δ strain revealed greater than 2-fold increase of gsf2+ and pfl3+ transcripts and a 60-fold increase of the SPAC1F8.02c transcript suggesting that these two flocculin genes and this uncharacterized glycoprotein gene may be responsible for the flocculent phenotype in this mutant. In contrast to the yox1Δ and sre2Δ mutants, mbx2+ did not show differential expression in the cbf11Δ strain compared to wild type. However, the cbf12+ transcript was upregulated 1.8-fold in the cbf11Δ strain. This suggests that cbf11+ may regulate flocculation through cbf12+, in agreement with previous reports of the antagonistic functions of cbf11+ and cbf12+ in this process [22].
We next determined whether the flocculation caused by the deletion of yox1+, sre2+ or cbf11+ was also dependent on gsf2+. The absence of gsf2+ was sufficient to abolish the flocculation in yox1Δ, sre2Δ, and cbf11Δ strains, even though gsf2+ was not always the most highly-expressed flocculin gene (Figure 7A and 7C, Table S14). Taken together, these results suggest that the expression of the dominant flocculin Gsf2 is responsible for the bulk of flocculation observed in yox1Δ, sre2Δ and cbf11Δ strains.
The role of flocculation by Adn2 and Adn3 is influenced by genes encoding cell wall–modifying enzymes and gsf2+
The transcription factor genes adn2+ and adn3+ are orthologous to Sacc. cerevisiae FLO8 (http://www.pombase.org/) and exhibit defects in invasive growth and cell-to-surface adhesion when deleted during nitrogen starvation [29]. From our screens, we discovered that the overexpression of adn2+ and adn3+ triggered minor flocculation while loss of adn2+ and adn3+ prevented flocculation in flocculation-inducing medium (Figure 8A and 8B, respectively). The flocculent phenotype of adn2OE and adn3OE strains was disrupted by the addition of galactose (data not shown). To identify the target genes of Adn2 and Adn3 that are involved in flocculation, expression microarray profiling was performed on nmt1-driven adn2OE and adn3OE strains (Tables S11 and S12). Surprisingly, gsf2+ transcript levels were relatively unchanged and the majority of pfl+ genes were downregulated in both overexpression strains (Figure 8C). Consistent with these results were the observations that mbx2+ and cbf12+ transcripts were downregulated greater than 2-fold in both adn2OE and adn3OE strains, whereas rfl1+ transcript levels were not differentially regulated (Figure 8C). Therefore, it appeared that the flocculent phenotype of adn2+ and adn3+ overexpression could not be attributed to the pfl+ genes identified in this study. These results led us to consider that perhaps the expression of other genes besides these encoding for flocculins could be responsible for triggering flocculation in adn2OE and adn3OE strains.
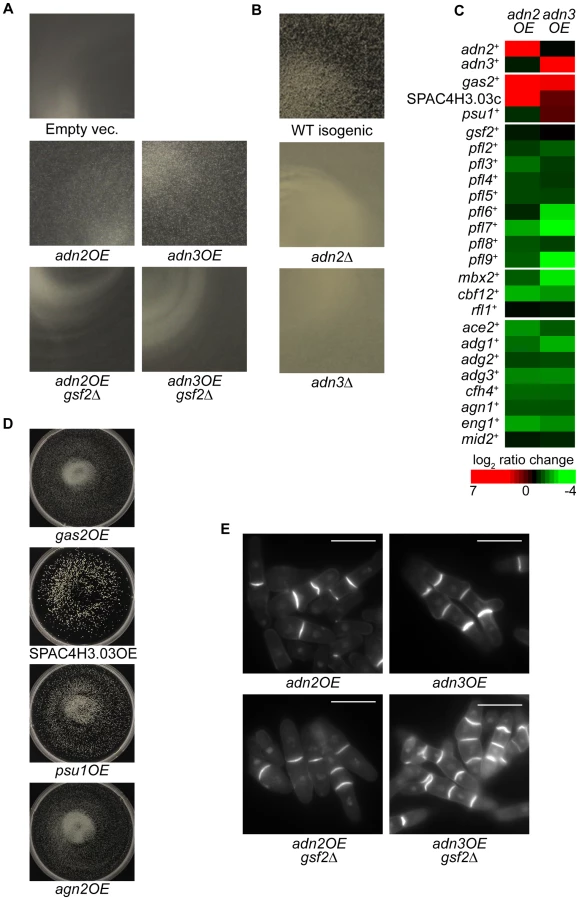
Interestingly, some of the aforementioned cell wall-remodeling enzymes (gas2+, psu1+ and SPAC4H3.03c) were also highly upregulated in both adn2OE and adn3OE strains (Figure 8C, Table S13). For example, gas2+ and SPAC4H3.03c were the highest induced genes in the adn2OE strain (17.9 and 36.8-fold, respectively) and also appeared within the top 20 most induced genes in the adn3OE strain. These genes were also induced in the mbx2OE strain except for psu1+ (Figure 2A). Overexpression analysis was subsequently carried out to determine if these genes possessed some role in flocculation. Although agn2+ was not upregulated in the adn2OE and adn3OE strains, it was included in the overexpression analysis because it was the second most induced gene (91-fold), as well as detected by ChIP-chip in the mbx2OE strain. Indeed, the single overexpression of these four genes resulted in flocculation after 5-days (including 3rd day sub-culturing into fresh medium) in medium lacking thiamine, implicating the involvement of these cell wall-remodeling enzymes in flocculation (Figure 8D, Table S14). Since deletion of adn2+ and adn3+ results in defects of invasive growth and cell-to-surface adhesion in response to nitrogen starvation, we wanted to determine if the single overexpression of gas2+, agn2+, psu1+ and SPAC4H3.03c could cause enhancement of these processes. We discovered that the single overexpression of these four cell wall-remodeling genes increased cell-to-surface adhesion, but not invasive growth relative to wild type under the nitrogen-deprivation condition (Figure S1). Because gsf2+ encodes the dominant flocculin, we also investigated whether the flocculation caused by adn2+ and adn3+ overexpression was dependent on gsf2+. Deletion of gsf2+ completely abrogated the flocculation in adn2OE and adn3OE strains (Figure 8A, Table S14).
In addition, the adn2OE and adn3OE strains exhibited cell separation defects such as the formation of multisepta and forkhead phenotypes (Figure 8E). The cell separation defect was more severe when adn3+ was overexpressed. We next determined whether the putative target genes involved in the flocculation of adn2OE and adn3OE strains also played a role in the multisepta phenotype. Overexpression of adn2+ and adn3+ in the gsf2Δ background did not alter the multisepta phenotype (Figure 8E), while the overexpression of gas2+, SPAC4H3.03c, psu1+ and agn2+ did not lead to formation of multisepta (data not shown). These results suggest that Adn2 and Adn3 may regulate cell separation and flocculation independently through different sets of target genes. Our microarray expression data suggests that Adn2 and Adn3 may control cell separation through ace2+, which encodes a major transcriptional activator of this process (Alonso-Nuñez et al., 2005). Overexpression of adn2+ and adn3+ resulted in the down-regulation of ace2+ and many of its known target genes such as adg1+, adg2+, adg3+, cfh4+, agn1+, eng1+, and mid2+ by 1.5 to 3.4-fold (Figure 8C).
In summary, the regulation of flocculation by adn2+ and adn3+ is likely mediated by the induction of genes encoding the cell wall-remodeling enzymes Gas2, SPAC3H3.03c, and Psu1. The regulation of these genes is independent from Mbx2 because mbx2+ was downregulated in the adn2OE and adn3OE strains. Although gsf2+ transcript level was not significantly upregulated by adn2+ and adn3+ overexpression, it was sufficient to abrogate the flocculation when deleted. However, it is possible that other cell surface glycoprotein genes not investigated in this study but were upregulated may also play a significant role in the flocculation function of adn2+ and adn3+.
Discussion
In this study, we have deciphered a significant portion of the transcriptional-regulatory network governing flocculation in S. pombe. To date, few transcription factors and their target genes that function in flocculation have been identified. The MADS box transcription factor Mbx2 positively regulates flocculation by induction of the flocculin gene gsf2+, while the CSL transcription factors Cbf11 and Cbf12 repress and activate flocculation, respectively, but their target genes are not known [21], [22]. We have substantially expanded our limited knowledge of the flocculation transcriptional-regulatory network by the identification of several novel transcriptional activators (Adn2 and Adn3) and repressors (Rfl1, Yox1 and Sre1), and their putative target genes that function in flocculation. In addition, novel target genes of Mbx2, Cbf11 and Cbf12 were identified. The putative target genes of the transcription factors implicated in flocculation encode for several cell surface glycoproteins (gsf2+ and pfl2+–pfl9+) and cell wall-remodeling enzymes (agn2+, psu1+, SPAC4H3.03c and gas2+). These target genes were sufficient to trigger flocculation when overexpressed. Moreover, instances of regulation between transcription factors (Rfl1 repression of mbx2+), as well as positive (mbx2+ and cbf12+) autoregulation were detected within the flocculation network.
Mbx2 and Rfl1 appeared to be the major positive and negative regulators of flocculation, respectively, based on the largest flocs observed in the mbx2OE and rfl1Δ strains compared to the other flocculent mutants in this study. Our initial efforts to identify the target genes of Mbx2 and Rfl1 revealed several putative flocculin genes that were strikingly upregulated in the mbx2OE and rfl1Δ flocculent mutants. Previously, Gsf2 was the only S. pombe flocculin demonstrated to be directly involved in flocculation, and its transcription was influenced by the activity of Mbx2 [20], [21]. Similar to these studies, we also found that overexpression of gsf2+ triggers flocculation while loss of gsf2+ abrogates the flocculent phenotype of several mutants including mbx2OE. Here, we identified an additional eight flocculin genes (pfl2+–pfl9+) as putative target genes of Mbx2. Seven of these target genes (pfl3+–pfl9+) were reported to contain tandem repeats found in fungal adhesins, while pfl2+ is a sequence orphan predicted to encode a GPI-anchored protein [23], [27]. Seven pfl+ genes (gsf2+/pfl1+, pfl3+, pfl4+ and pfl6+–pfl9+) have been reported to be upregulated in loss-of-function flocculent mutants of Cdk8 module genes (cdk8+/srb10+, med12+/srb8+) suggesting that the transcriptional repression of these putative flocculin genes may be controlled by Mediator [24]. The transcriptional repression of flocculin genes by Mediator may not be direct, but could be through mbx2+ since its expression is highly upregulated in the cdk8 kinase-mutant and med12Δ strain (9 and 13-fold increase, respectively, within top 11 up-regulated genes, found in supplementary data [24]). This proposed role of Mediator appears conserved in Sacc. cerevisiae as FLO genes are similarly upregulated in cdk8 mutants [38]. Despite these observations, no direct evidence has been shown aside from the gsf2+ study by Matsuzawa et al. [20] that the pfl+ gene products are actually flocculins. We have shown that this is indeed the case as single and double overexpression of the pfl+ genes is sufficient to trigger flocculation and that this flocculation is galactose-specific.
The degree of flocculation triggered by single overexpression of the pfl+ genes varied, with gsf2+ and pfl9+ producing the largest and smallest flocs, respectively (the pfl numbers correspond roughly to the degree of flocculation upon overexpression). This result indicates that gsf2+ encodes the most dominant flocculin compared to the other pfl+ genes. In agreement are the observations that only deletion of gsf2+ and not the other pfl+ genes prevented flocculation in flocculation-inducing medium, and reduced the constitutive flocculent phenotype to the greatest extent of all the transcription factor mutants examined in this study. Moreover, the strength of the flocculins was directly correlated with the amount of reduction in mbx2OE flocculation observed in the various pfl deletion backgrounds (Figure 2B). These observations are similar in Sacc. cerevisiae, where overexpression of FLO1 produces the strongest flocculation compared to FLO5, FLO9, FLO10 and FLO11 [8],[12]. Furthermore, the flocculation mediated by pfl+ genes was additive as observed in our double deletion and co-overexpression experiments (Figure 2B and Figure 3B). These results suggest that the varying strengths of flocculation exhibited by S. pombe strains could be attributed to the upregulation of different combinations of pfl+ genes.
We identified Rfl1, an uncharacterized Zn(2)-Cys(6) transcription factor as a novel repressor of flocculation in fission yeast. The repression of flocculation by Rfl1 appears to be primarily mediated by the inhibition of gsf2+ expression since loss of gsf2+ can abrogate the constitutive flocculent phenotype of the rfl1Δ mutant. Rfl1 represses gsf2+ either directly by association with its promoter or indirectly by inhibition of mbx2+ transcription, thereby forming an inhibitory feed-forward loop (coherent type 2) within the transcriptional-regulatory network (Figure 9). These results indicate that Mbx2 and Rfl1 are opposing transcription factors, and the latter inhibits mbx2+ and gsf2+ expression under non-inducing conditions of flocculation.

Aside from its role in flocculation, Rfl1 may have a role in regulating genes involved in carbohydrate metabolism such as glycolysis and gluconeogenesis. Rfl1 appeared to be associated with promoters of genes enriched in glucose catabolic and metabolic processes (p-values = 0.00092 and 0.00269, respectively) including adh1+, hxk2+, pfk1+, tpi1+, adh4+, pgi1+, gpd3+, tdh1+, pgk1+, fba1+, eno101+, pyr1+, SPCC794.01c (predicted glucose-6-phosphate 1-dehydrogenase), SPBC2G5.05 (predicted transketolase) and SPBC660.16 (phosphogluconate dehydrogenase). Most of these genes with the exception of fba1+, eno101+, SPCC794.01, and SPBC660.16 were upregulated 1.2 to 26-fold in the rfl1Δ strain (Table S4). From these data, we speculate that Rfl1 could serve as a negative transcriptional regulator of several enzymes involved in the glycolysis and gluconeogenesis. Because flocculation and invasive growth are associated with nutritional limitation, Rfl1 may coordinate the expression of genes involved in flocculation and carbohydrate metabolism in fission yeast.
Previously, the CSL proteins Cbf11 and Cbf12 were shown to exhibit antagonistic roles in flocculation [39]. Overexpression of cbf12+ or loss of cbf11+ triggers flocculation. However, none of their target genes have been identified. We present supportive evidence that Cbf12 induces flocculation by directly activating the transcription of gsf2+. In addition, gsf2+ expression is up-regulated approximately 2.4-fold in the cbf11Δ strain suggesting that the repressive flocculation function of Cbf11 may also be directly mediated through gsf2+. The activation and repression of gsf2+ transcription by Cbf12 and Cbf11, respectively, may occur by competitive binding to promoter sites since both transcription factors have been shown to interact with a canonical CSL consensus sequence in vitro [39]. Several putative sites with six out of seven nucleotide match to the canonical CSL consensus sequence are located within 900 base pairs of the gsf2+ promoter (data not shown). Further experimentation would be required to verify this proposed mechanism of gsf2+ transcriptional regulation by Cbf11 and Cbf12. It is likely that cbf12+ plays a lesser role in activating flocculation compared to mbx2+ since the floc size resulting from cbf12+ overexpression is considerably smaller than the mbx2OE strain. Also unlike mbx2+, deletion of cbf12+ is not sufficient to abrogate the flocculation of the rfl1Δ strain (data not shown). These data suggest that the flocculent phenotype of the cbf12Δ rfl1Δ double mutant is probably caused by the presence of mbx2+ activity.
CSL transcription factors are components of the conserved Notch signaling pathways in metazoans which primarily function in cell-to-cell communication during development [40]. Although multiple fungal CSL proteins have been discovered, their exact roles remain unclear in unicellular organisms [39]. Flocculation has been described as a manifestation of social behaviour in yeast with a purpose of enhancing survival under stressful conditions [3]. Therefore, it is conceivable that CSL transcription factors originated as regulators of this primitive form of cell-to-cell communication, and later evolved into the metazoan Notch signaling pathway.
We also discovered novel functions of the Yox1 and Sre2 transcription factors in the repression of flocculation. Loss of yox1+ or sre2+ results in a mild flocculent phenotype. The Yox1 homeodomain transcription factor functions as a repressor of MBF (Mlu1 binding factor) target genes to prevent their inappropriate expression at the end of S-phase [36]. Transcriptional repression of MBF target genes is mediated by the direct interaction of Yox1 and Nrm1 to the MBF complex [41]. Deletion of yox1+ causes a cell cycle delay and results in elevated constitutive expression of MBF gene targets [36]. Similarly, these genes (e.g. cdc18+, cdc22+, cdc10+, cdt1+, cdt2+, cig2+ and nrm1+) were also found to be upregulated 2.2 to 6.4-fold in our yox1Δ microarray expression data (Table S8). We found that the flocculent phenotype of the yox1Δ strain is also dependent upon gsf2+. However, the pfl+ genes including gsf2+ were not highly expressed in the yox1Δ strain. One possible explanation why pfl1+ genes were not highly expressed in the yox1Δ strain is that our experiments were performed under asynchronous culturing conditions, and therefore, the upregulation of pfl+ genes including gsf2+ could have been obscured if their expressions were periodically controlled in the vegetative cell cycle. However, it is unlikely that Yox1 regulates the pfl+ genes directly because previous chIP-chip analysis did not detect binding of Yox1 to the promoters of pfl+ genes [36]. Although there has been solid evidence linking yeast morphogenesis events such as pseudohyphal and hyphal growth to cell cycle regulators [42]–[47], the relationship between cell cycle control and flocculation remains unclear. A flocculation function for yox1+ has not been reported in other yeasts. However, disruption of YOX1 in the Sacc. cerevisiae ∑1278b strain inhibited filamentous invasive growth, a process usually associated with flocculation during nutritional limitation, while deletion of C. albicans NRM1 reduced flocculation [48], [49].
Sre2 is an uncharacterized membrane-tethered helix-loop-helix transcription factor predicted to be an ortholog of mammalian SREBP-1a, which is responsible for the transcriptional activation of genes needed for uptake and synthesis of cholesterol, fatty acids, triglycerides, and phospholipids [50]. While sre1+, a paralog of sre2+ has been shown to function in the transcriptional activation of sterol-biosynthetic and hypoxic-adaptation genes, there has been no direct evidence that sre2+ plays similar biological roles [37]. Loss of sre2+ results in the upregulation of gsf2+, pfl3+ and fta5+ transcripts (3.76, 2.51 and 3.08-fold, respectively) (Figure 7C, Table S9) which may contribute to its flocculent phenotype. The sre2Δ flocculent phenotype requires gsf2+ activity and is independent of mbx2+ and cbf12+ since these transcripts are downregulated in the deletion mutant.
In addition, the microarray expression profiles of yox1Δ and sre2Δ strains displayed similar differential gene expression despite the supposedly different functions of these transcription factors (Figure 7B). Mitochondrial genes were found to be highly upregulated in both deletion mutants (Tables S8 and S9). This occurrence may not be unexpected for Sre2 if it has a similar role in hypoxia as Sre1 where mitochondrial function is probably impaired [37]. It is currently not clear whether the mitochondrial genes are direct targets of Yox1 and Sre2 or induced in response to an altered physiological state in the deletion mutants. Interestingly, mitochondrial activity has been reported to be important for flocculation and invasive growth in Sacc. cerevisiae [48], [51]. Disruption of mitochondrial activity has been shown to alter the synthesis and structure of the cell wall, possibly by interfering with the interactions of flocculins and their substrates [52]. Based on these observations, the flocculent phenotype of yox1Δ and sre2Δ strains could be partially the result of enhanced mitochondrial activity from the upregulation of mitochondrial genes.
A genome-wide systematic deletion screen previously uncovered a cell-to-surface adhesion function that is sensitive to the presence of galactose for the Adn2 and Adn3 transcription factors [29]. Here, we discovered that adn2+ and adn3+ have additional functions in flocculation. Overexpression of adn2+ and adn3+ induced minor flocculation while loss of these genes prevented flocculation in inducing glycerol medium (Figure 8A and 8B). However, the flocculent phenotype of the adn2OE and adn3OE strains appeared to be primarily caused by the differential regulation of genes encoding cell wall-remodeling enzymes rather than flocculins. Several genes encoding cell wall-remodeling enzymes (gas2+, agn2+, psu1+ and SPAC4H3.03c) were highly induced when mbx2+, adn2+ or adn3+ was overexpressed. In the adn2OE strain, gas2+ and SPAC4H3.03c were the most highly induced genes (17.9 fold and 36.8-fold, respectively) (Figure 8C, Table S11) while in the adn3OE strain, these two genes and psu1+ appeared within the top 20 up-regulated genes (Table S12). Similarly, in the mbx2OE strain, gas2+, agn2+, and SPAC4H3.03c appeared within the top 100 up-regulated genes (greater than 3.7-fold increase, Figure 2A). We found that the single overexpression of these four genes could trigger flocculation (Figure 8D). Cell wall remodeling is an essential process for proper growth and adaptation to environmental stresses in yeast cells. Part of the cell wall-remodeling process involves the dissolution of sugar moieties in the glucan layer and elongation of glucan chains by glycoside hydrolases and glycosyltransferases, respectively. Among these four genes, three (agn2+, psu1+ and SPAC4H3.03c) encode for glycoside hydrolases while the fourth (gas2+) encodes for a glycosyltransferase. Agn2 is an endo-(1,3)-α-glucanase that hydrolyzes (1,3)-α-glucans of the ascus wall for ascospore release [53], [54]. Although agn2+ function appears only specific for sporulation, its ectopic expression could alter the cell wall structure during vegetative growth by inappropriate hydrolysis of (1,3)-α-glucan. Similarly, inappropriate glucan hydrolysis of the cell wall could be occurring as a result of ectopic expression of SPAC4H3.03c which encodes a putative (1,4)-α-glucanase (Hertz-Fowler et al., 2004). Psu1, which exhibits close homology to the members of the SUN family in Sacc. cerevisiae and C. albicans, as well as the BglA beta glucosidase of C. wickerhamii, has an essential function in cell wall synthesis [55]. Loss of psu1+ activity conferred resistance to (1,3)-β-glucanase suggesting that Psu1 may influence the amount or structure of (1,3)-β-glucan in the cell wall [55]. In addition, the (1,3)-β-glucanosyltransferase Gas2 has been shown to lengthen glucan chains during cell wall assembly and its overproduction is able to suppress the cell wall defect and lethality of gas1Δ cells [56]. Then how does the overexpression of these cell wall-remodeling genes trigger flocculation in S. pombe cells? The expression of flocculin genes during vegetative growth is not well characterized in yeasts, but studies in Sacc. cerevisiae indicate that flocculin synthesis and insertion into the cell wall initiate in early exponential phase prior to the onset of flocculation during stationary phase [57]. This suggests that the flocculins are already present in the cell wall, but cannot induce flocculation because of inaccessibility to cell surface oligosaccharides. We speculate that the restructuring of the β-glucan layer during cell wall remodeling may result in the rearrangement of flocculins that enhances galactose oligosaccharide binding, thereby promoting flocculation. Several lines of evidence in Sacc. cerevisiae support this hypothesis. First, alteration of cell wall structure by disruption of PKC1 activity results in flocculation [58]. Second, heat shock induces flocculation and regulation of cell wall-remodeling genes via the Hsf1 transcription factor [57], [59]. Currently, we cannot rule out that agn2+, psu1+, SPAC4H3.03c and gas2+ are the only cell wall remodeling enzymes that can trigger flocculation when overexpressed. Other genes with potential functions in cell wall modification and integrity such as gas4+, gma12+, meu7+, agl1+, meu10+ and mde5+ were also detected as putative target genes of Mbx2 (Table S2). In contrast, there was little change in gsf2+ transcript levels in the adn2OE or adn3OE strains compared to the empty vector control. However, the flocculation triggered by adn2+ and adn3+ overexpression was abrogated in a gsf2Δ background indicating that gsf2+ was indispensible for this process (Figure 8A). Altogether, these results suggest that Gsf2 is likely expressed in the cell wall as an inactive flocculin, and the cell wall remodeling resulting from adn2+ and adn3+ overexpression alters the arrangement of Gsf2 and possibly other flocculins that now becomes favorable for flocculation.
The single overexpression of the cell wall-remodeling genes triggered flocculation to a greater extent than the adn2OE and adn3OE strains. A possible explanation for the different degrees of flocculation between the transcription factor and its target genes could be that overexpression of adn2+ and adn3+ causes reduced fitness due to toxicity effects associated with a greater misregulation of genes compared to the aberrant production of a single enzyme. Consistent with this theory is that adn2OE and adn3OE strains exhibited additional phenotypes including septation defects (Figure 8E) which were not observed when gas2+, agn2+, psu1+ and SPAC4H3.03c were overexpressed (data not shown). Furthermore, a systematic overexpression analysis of 5280 genes in Sacc. cerevisiae revealed that genes encoding for transcription factors, signalling molecules and cell cycle regulators were more likely to cause reduced fitness [60].
In S. pombe, cell separation involves the transcriptional activation of adg1+, adg2+, adg3+, agn1+, eng1+, cfh4+ and mid2+ by the Ace2 transcription factor, which is in turn regulated by the Sep1 forkhead transcription factor [61]–[64]. We discovered that the adn2OE or adn3OE strains displayed multisepta and forkhead phenotypes similar to loss-of-function mutations of these cell separation genes. The cell separation defect in adn2OE and adn3OE strains is likely due to the downregulation of ace2+ transcription since ace2+ and its target genes were substantially downregulated in these strains (Figure 8C). However, sep1+ transcript levels remained unchanged in the adn2OE and adn3OE strains indicating that their involvement in cell separation phenotype could be either downstream of sep1+ or parallel to the sep1+ pathway. In additional to its flocculation role, Adn2 and Adn3 appear to have a separate function in cell separation perhaps by directly or indirectly repressing ace2+ transcription. Experiments are planned in the future to address these possibilities.
Interestingly, we also found some evidence that supports a role of Mbx2 and Cbf12 in cell separation perhaps through repression of ace2+ activity. Overexpression of mbx2+ and cbf12+ results in significant down-regulation of all seven Ace2 target genes approximately 1.5 to 3.4-fold relative to the empty vector control. (Tables S2 and S6). The mbx2OE strain indeed showed septation defects but were slightly different in nature than the adn2OE and adn3OE strains with less multi-septation and more mislocalization of septum material (data not shown). Moreover, overexpression of cbf12+ has been reported to produce multisepta phenotypes albeit at a low frequency [39]. These observations indicate the possible existence of crosstalk between flocculation and cell separation pathways mediated by the Mbx2, Cbf12, Adn2 and Adn3 transcription factors (Figure 9).
A comparison between the flocculation network of budding and fission yeast revealed both conserved and divergent features within the transcriptional circuitry. In Sacc. cerevisiae, the positive and negative transcriptional controls of the dominant flocculin gene FLO1 by Flo8p or Mss11p, and Sfl1p, respectively, draw parallel to gsf2+ regulation by the Mbx2-Rfl1 and Cbf12-Cbf11 opposing transcription factors in S. pombe [16], [19]. The conservation of the flocculin genes between these two yeasts is apparent among these transcription factors. Similar to Mbx2, Mss11p and Flo8p appear to activate multiple flocculin genes (FLO1, FLO9 and FLO11), while the latter may also regulate genes encoding cell wall enzymes (STA1 and SGA1 which both encode glycoside hydrolases) [9], [16]. The putative target genes of the Sfl1 repressor have been reported to include FLO1 and FLO11 [19], [65]. In contrast to the conservation of the flocculin genes, the types of transcription factors involved in flocculation are quite different between the two yeasts. Mbx2 belongs to the MADS box family, while the DNA-binding domains of Flo8p and Mss11p have not been defined. In addition, the Sfl1p and Rfl1 repressors contain the heat shock factor and Zn(2)-Cys(6) DNA-binding domains, respectively. Moreover, CSL transcription factors (Cbf11/Cbf12) are not found in Sacc. cerevisiae. These observations would imply that the cis-regulatory elements controlling transcription of the flocculin genes have likely undergone considerable rewiring within the transcriptional-regulatory network between the two yeasts. However, it was recently demonstrated that heterologous expression of Flo8p and Mbx2 could induce gsf2+ and FLO1 transcription in fission and budding yeast, respectively [20], [21]. Therefore, despite the divergent types of transcription factors controlling flocculin gene expression in the two yeasts, there may be some degree of conservation among the cis-regulatory sequences.
Although the transcription factors regulating flocculation appear to be quite different between the two yeasts, the downstream transcriptional events involved in the repression of flocculin genes are likely to be conserved. Disruption of genes encoding the Ssn6p-Tup1p general corepressor or the Cdk8 module of Srb/Mediator complex have been shown to cause upregulation of flocculin genes and constitutive flocculation in S. pombe and Sacc. cerevisiae [65]. In the latter yeast, Sfl1p represses FLO1 and FLO11 transcription through physical interactions with Ssn6p and Srb proteins (Srb8p, Srb9p and Srb11p) [19], [65], [66]. Moreover, Sfl1p has been reported to repress FLO8 in Saccharomyces diastaticus. The observation that Sfl1p can repress FLO1 transcription directly and indirectly through FLO8 seems very similar to the inhibitory feed-forward regulation of gsf2+ by Rfl1 in S. pombe. If these connections are truly analogous, then there is a possibility that Rfl1 repression could also be mediated through physical interactions with the Cdk8 module proteins. In the srb10− mutant, gsf2+ and mbx2+ expression are upregulated suggesting that its flocculent phenotype could be caused by a failure to repress mbx2+ transcription [24]. In addition, the flocculent phenotype of tup11+/tup12+ mutants [25] and the abrogation of lkh1Δ flocculation in the absence of mbx2+ [21] supports the role of Tup11/12 corepressor in Mbx2-Rfl1-mediated flocculation. Taken together, we speculate that Srb10 and Tup11/12 activity and binding may be required for Rfl1-mediated repression of gsf2+ and mbx2+. Future experiments focusing on the interactions between Rfl1, Tup11/12 and Srb8-10 in relation to flocculation would provide clarification to our speculation.
Our analyses of the transcription factors implicated in flocculation of S. pombe revealed the possible existence of several network motifs including positive autoregulation of mbx2+ and cbf12+ and regulation of gsf2+ by a inhibitory feed-forward loop (coherent type 2). The latter involves the Rfl1 transcriptional repression of gsf2+ directly and indirectly by inhibition of mbx2+ expression. Autoregulatory motifs have not been detected so far for FLO8, MSS11 and SFL1. The discovery of these network motifs in S. pombe suggests that the transcriptional inhibition of gsf2+ could occur more rapidly than its transcriptional activation. Experimental and modeling studies have proposed that positive and negative autoregulation of transcription factors generate slow and fast response times, respectively, within a transcriptional-regulatory network [67]. Under positive autoregulation, the synthesis rate of the transcription factor is initially slow at low concentrations, but increases as the concentration of the transcription factor reaches the activation threshold of the promoter, while negative autoregulation accelerates the attainment of steady state levels of the transcription factor [67]. Moreover, the inhibitory feed-forward motif of Rfl1 seems to indicate that repression of gsf2+ expression likely happens in a shorter period compared to its activation. Altogether, these data suggest that the onset of flocculation may occur gradually while repression of the flocculation pathway is a much faster process. Consistent with this speculation is the observation that it requires several days for wild-type S. pombe cells to undergo flocculation when grown in inducing medium.
In summary, we have provided an initial and substantial view of the transcriptional-regulatory network governing flocculation in S. pombe. Found within this network are the master regulators Mbx2, Cbf12, Adn2 and Adn3, which are able to trigger flocculation when overexpressed by the activation of their target genes encoding for flocculins and cell wall-remodeling enzymes. In addition, several repressors including Rfl1 were uncovered that play a major role in the regulation of these target genes. However, significant gaps of knowledge surrounding the transcriptional-regulatory network still remain. The environmental cues that impinge upon the activity of the positive and negative regulators, as well as the dynamics of transcription factor binding and regulation of target genes during the onset of flocculation remain to be elucidated. Also, although gsf2+ encodes the dominant flocculin, it is currently unclear whether the other flocculins have nonessential or more specialized roles during flocculation. Detailed analyses of the temporal and spatial expression of the pfl+ genes would be required to address these questions. Moreover, the exact mechanism of how other biological processes such as cell wall restructuring and mitochondrial function influence flocculation is unknown. Further studies to expand our knowledge of this transcriptional-regulatory network would provide a more comprehensive understanding of flocculation control and contribute to a valuable resource for the improvement of industrial yeast applications.
Materials and Methods
Yeast strains, media, and general methods
All strains used in this study are listed in Table S1 and were maintained on YES or EMM medium. Geneticin, nourseothricin, and thiamine hydrochloride were added to media at a concentration of 150 mg/L, 100 mg/L, and 15 µM, respectively. EMM medium was supplemented with amino acids when necessary at 225 mg/L each for adenine, leucine, and uracil. Matings were performed on SPAS medium. Wild type and deletion strains were assayed for flocculation in YEGlyEtOH (flocculation-inducing) medium containing 1% (w/v) yeast extract, 3% (v/v) glycerol, and 4% (v/v) ethanol. Overexpression strains containing ORFs under control of the nmt1 or nmt41 promoter were grown in EMM minus thiamine medium. Standard genetics and molecular biology techniques were performed as described in [68].
Construction of deletion and GFP-tagged strains
A PCR-based stitching method was utilized to construct the deletion and epitope-tagged strains. For construction of deletion strains, ∼500 bp fragments upstream and downstream of the ORF and the KanMX6 or NatMX6 cassette were PCR-amplified and gel-purified. The 3′ end of the upstream fragment and 5′ end of the downstream fragment contained ∼25 bp homology to the selectable marker cassette sequence. Approximately equimolar amounts (∼40 ng) of each PCR fragment were combined and stitched together in a 20 µl PCR reaction (0.2 mM dNTPs and 0.4 units of Phusion HF DNA polymerase (New England Biolabs), and subjected to one cycle of 98°C (30 sec), 5 cycles of 98°C (15 sec), 60°C (1 min), and 72°C (1–2 min) and a final extension at 72°C (5 min). The stitched product was then amplified in a 50 µl PCR reaction by combining the entire stitched reaction with 6 nmol dNTPs, 0.6 units of Phusion HF DNA polymerase and 20 pmol each of the outer pair of primers and then subjected to one cycle of 98°C (30 sec), 30 cycles of 98°C (10 sec), 60°C (30 sec) and 72°C (2 min), and a final extension at 72°C (5 min). The amplified product was gel-purified and transformed into the appropriate strain by lithium acetate transformation. A similar strategy was used to construct GFP-tagged transcription factors under the control of the native promoter. To tag the transcription factor with GFP at the C-terminus, ∼500 bp upstream and downstream fragments flanking the stop codon and the GFP-KanMX6 cassette (amplified from pYM27 plasmid, [69]) were PCR-amplified for the stitching reaction as described above. To conserve the native promoter in the N-terminal GFP fusion of Mbx2, 1 kb upstream of the mbx2+ start codon was amplified along with four other fragments for PCR stitching: (1) ∼500 bp upstream of the aforementioned 1 kb fragment; (2) ∼500 bp downstream of the mbx2+ start codon; (3) KanMX6 cassette and; (4) the GFP ORF with its stop codon removed and a GDGAGL linker added (adapted from [70]). All five fragments contained ∼25 bp overlapping homology to their respective flanking fragments and were PCR-stitched as described above. Proper gene deletion and GFP tagging were confirmed by colony PCR screen and the resulting amplicons sequenced.
Construction of overexpression strains
Genes were overexpressed with the nmt1 promoter by cloning the entire ORFs of interest into the pREP1 or pREP2 vector. For ChIP-chip experiments, C-terminal triple HA-tagged Mbx2, Rfl1, and Cbf12 were expressed with the nmt41 promoter by cloning the corresponding ORFs into pSLF272 [71]. All the clones were PCR-confirmed, sequenced, and transformed into appropriate strains by the lithium acetate method. Expression of the HA-tagged proteins was verified by western blotting with anti-HA F-7 antibody (Santa Cruz Biotechnology, Santa Cruz, CA).
Microarray expression profiling
Strains overexpressing the triple HA-tagged Mbx2, Rfl1, and Cbf12 were grown in 200 ml of EMM medium containing appropriate supplements without thiamine for 18-20 hr to induce the nmt41 promoter. The empty vector control strain was cultured concurrently to a matching cell density of ∼8×106 cells/ml prior to harvesting. The experimental culture was divided into two, each for ChIP-chip and microarray expression profiling while the control culture was only utilized in the latter. The expression profiling cultures were harvested by centrifugation (1800× g, 3 min, 20°C), followed by immediate freezing of the cell pellets in liquid nitrogen. Culturing of adn2OE and adn3OE strains were performed similarly except that these genes were driven by the nmt1 promoter and were not epitope-tagged. For transcription factor deletion strains (rfl1Δ, cbf11Δ, sre2Δ, and yox1Δ), the mutant and an isogenic wild-type strain were concurrently grown in YES medium and harvested at a similar cell density as described above. Total RNA extraction, mRNA isolation, reverse transcription with aminoallyl-dUTP (Sigma-Aldrich, Oakville, ON), and Cy™3/Cy™5 (GE Healthcare, Buckinghamshire, UK) dye coupling of cDNA were performed with dye reversal as previously described [72]. Purified Cy™3 - and Cy™5-labelled cDNA (1 µg in total) was hybridized onto custom-designed 8×15 K Agilent expression microarrays containing 60mer probes to all S. pombe ORFs in 2–3 times coverage per gene. The hybridization procedure was carried out according to the manufacturer's instructions (Agilent Technology, Santa Clara, CA) with the exception for the use of Human Cot-1 DNA. The microarrays were washed in 6× SSPE/0.005% sodium N-lauroylsarcosine at room temperature for 5 min followed by a second wash in pre-heated 42°C 0.6× SSPE for 2 min.
The microarrays were scanned with a GenePix4200A scanner (Molecular Devices, Sunnyvale, CA). The raw microarray data was lowess normalized [73] and the average log2 ratios with the corresponding t-test p values [74] from the dye-swap experiments were obtained using the R Bioconductor Limma package. Heat map images of the microarray expression and ChIP-chip data were constructed with Cluster 3.0 [75] and Java Treeview 1.1.6r2 [76]. The microarray expression data has been submitted to the NCBI Gene Expression Omnibus Database (GSE41730).
ChIP–chip experiments and data analysis
Culturing of the HA-tagged transcription factor strains are described above. The culture was fixed by the addition of a final concentration of 1% formaldehyde and agitation for 30 min at room temperature. The formaldehyde was quenched by the addition of 2.5 M glycine to a final concentration of 125 mM and agitation for 5 min at room temperature. The cells were then centrifuged (800× g, 5 min, 4°C), washed twice in 25 ml 1× ice-cold PBS (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4 pH 7.4) and washed once with 2 ml ice-cold lysis buffer (50 mM NaCl, 50 mM HEPES-KOH pH 7.5, 0.1% SDS, 1% Triton X-100, 1 mM EDTA, 0.1% sodium deoxycholate and 1 tablet/50 ml Protease Inhibitor Cocktail (Roche Applied Science, Indianapolis, IN)). The cell pellet was resuspended in 1.6 ml lysis buffer and stored at −80°C.
The cell suspension was transferred to two 2 ml bead beating vials containing 800 µl of 0.5 mm Zirconia/Silica beads (BioSpec Products, Bartlesville, OK) and subjected to 3 cycles of alternating 2 min beating and 2 min incubation on ice with a Mini Beadbeater 16 (BioSpec Products, Bartlesville, OK). The lysed cells were collected by puncturing the bottom of the bead-beating vial with a flame-heated inoculating needle and placing the vial on a sonication tube nested in 10 ml disposable culture tubes prior to centrifugation (800× g, 3 min, 4°C). The cell pellet was resuspended, transferred to chilled microcentrifuge tubes, centrifuged (16,000× g, 15 min, 4°C) to remove unbound soluble proteins, and the resulting pellet resuspended in 800 µl of fresh lysis buffer in a sonication tube. Total cell lysate volume was adjusted to 2.2 ml with lysis buffer and subjected to 4 cycles of sonication and 1 min on ice incubation at 30% amplitude, 30 sec setting using a Sonic Dismembrator with a 1/8 tapered microtip probe (Thermo Scientific, Waltham, MA). The sonicated cell lysate was centrifuged (4600× g, 2 min, 4°C) and the supernatant stored at −80°C. The supernatant was tested to ensure that greater than 90% of the sonicated DNA was in the size range of 100 bp–1 kb by subjecting a sample (∼50 µl) of the supernatant to overnight reverse-crosslinking at 65°C and phenol-chloroform extraction, followed by gel electrophoresis of 3–5 µg of DNA. To immunoprecipitate the chromatin-bound transcription factor, 100–200 µl of Dynabeads conjugated with sheep anti-mouse IgG (Invitrogen Life Technologies, Carlsbad, CA) were washed twice in 800 µl ice cold 1× PBS-BSA (5 mg/ml BSA, 1× PBS), resuspended in 800 µl cold 1× PBS-BSA with 5 µg of anti-HA F-7 antibody (Santa Cruz Biotechnology, Santa Cruz, CA) and shaken gently for 2 hr at 4°C on a Labquake Tube Shaker (Thermo Scientific, Waltham, MA). The beads were washed twice in 1 ml cold deoxycholate buffer (100 mM Tris-HCl pH 8, 1 mM EDTA, 0.5% (w/v) sodium deoxycholate, 0.5% (v/v) NP-40, 250 mM LiCl) and twice in 1 ml cold lysis buffer. The beads were resuspended in 200 µl 1× PBS-BSA, combined with 400 µl of sonicated cell lysate, and shaken gently for 2 hr at 4°C. Four washes of 5 minutes each were next carried out: (1) 1.4 ml cold lysis buffer at 4°C; (2) 1.4 ml cold lysis buffer with 400 mM NaCl at 4°C; (3) 1.4 ml deoxycholate buffer at room temperature and; (4) 1.4 ml TE (pH 8) at room temperature. The transcription factor and bound DNA were eluted twice from the Dynabeads by incubating with 250 µl TES each (TE pH 8, 1% (w/v) SDS) at 65°C for 6 min. Dynabead washing and the supernatant collection were performed using DynaMag™−2 (Invitrogen, Carlsbad, CA). For the input DNA, 200 µl of the cell lysate was added to 300 µl TES. Both the immunoprecipitated and input cell lysates were incubated at 65°C overnight to reverse the DNA-protein cross-linking. Western blotting with anti-HA antibody was performed to confirm proper pull-down of the transcription factor.
For protein removal, both immunoprecipitated and input samples were incubated with 200 µg Proteinase K (Promega, Madison, WI) and 20 µg glycogen (Roche Applied Science, Indianapolis, IN) at 56°C for 2 hr. The DNA was then extracted by phenol-chloroform extraction, ethanol-precipitated overnight, washed once with 70% EtOH, resuspended in 42 µl TE containing 0.1 µg DNAse-free RNaseA (Roche Applied Science, Indianapolis, IN), and incubated for 30 min at 37°C.
Blunt ends were generated in the entire immunoprecipitate and input DNA samples with 1 unit of T4 DNA Polymerase (Invitrogen Life Technologies, Carlsbad, CA), 1× NEB Buffer #2 (New England Biolabs, Ipswich, MA), 5 µg NEB BSA, and 10 nmol dNTPs in a 110 µl reaction by incubation at 12°C for 20 min, followed by phenol-chloroform extraction and ethanol precipitation with 10 µg glycogen and 1/10 volume 3 M NaOAc. The DNA pellets were washed in 70% EtOH and resuspended in 25 µl water. Approximately 1/5 of precipitated input DNA was used in the subsequent ligation reaction as input DNA concentration was >100 times greater than that of immunoprecipitated DNA. For ligation of linkers to blunt ends, the resuspended DNA was incubated with 1000 units of concentrated T4 DNA Ligase (New England Biolabs, Ipswich, MA), 1× T4 DNA Ligase Buffer (Invitrogen Life Technologies, Carlsbad, CA), and 200 pmol annealed linker (15 µM Oligo #1 5′-GCGGTGACCCGGGAGATCTGAATTC-3′ and 15 µM Oligo #2 5′-GAATTCAGATC-3′ in 250 mM Tris) at 16°C overnight. The annealed linker and the ligation mix were kept on ice at all times prior to overnight incubation. The DNA was ethanol-precipitated, washed, and resuspended in 25 µl water as described above.
The ligated DNA was PCR-amplified by adding 15 µl of labeling mix (2 µl aa-dUTP dNTP mix containing 5 mM each dATP, dCTP and dGTP, 3 mM dTTP, and 2 mM aminoallyl-dUTP (Sigma-Aldrich, St. Louis, MO)), 1.25 µl 40 µM Oligo #1 (5′-GCGGTGACCCGGGAGATCTGAATTC-3′), 4 µl 10× ThermoPol Buffer (New England Biolabs, Ipswich, MA) and 7.75 µl water) in a PCR cycler paused at 55°C. A 10 µl enzyme mix containing 5 units of GoTaq DNA polymerase (Promega, Madison, WI), 0.001 units of Pfu Turbo DNA polymerase (Stratagene, La Jolla, CA), and 1× ThermoPol Buffer (New England Biolabs, Ipswich, MA) was added and the PCR proceeded with one cycle of 55°C (4 min); 72°C (5 min); 95°C (2 min) and 30 cycles of 95°C (30 sec); 55°C (30 sec); 72°C (1 min), followed by a final extension at 72°C (4 min). The PCR products were purified (QIAGEN, Valencia, CA) with a few modifications: (1) buffer PE was replaced with phosphate wash buffer (5 mM KPO4 pH 8.5, 80% ethanol) and (2) buffer EB was replaced with phosphate elution buffer (4 mM KPO4 pH 8.5). A sample of the purified PCR product was run on an agarose gel to check for fragment sizes ranging between 100 bp and 1 kb. The purified PCR products were quantified, and equal amounts of immunoprecipitated samples and corresponding input samples were coupled to Cy™3 and Cy™5 dyes as described above.
The labelled samples (total amount of 3–5 µg) were hybridized onto an Agilent 4×44 K S. pombe Genome ChIP-on-chip microarray according to the manufacturer′s instructions (Agilent Technology, Santa Clara, CA) except for the use of Human Cot-1 DNA. The washing and scanning of the microarrays were performed as described above. The ChIP-chip data was normalized by scaling in Limma [73] and analyzed by ChIPOTle Peak Finder Excel Macro [77] with the default setting of log2 ratio cut-off of 1. Peaks located within 3 kb upstream of a start codon and 2 kb downstream of a start codon within a coding region or 3′-UTR, in the case of short ORFs, were assigned to the gene. ChIP-chip data sets are found in Tables S3, S5, and S7. Genes with multiple peaks are noted in the data set with the peak values. The ChIP-chip data has been submitted to the NCBI Gene Expression Omnibus Database (GSE41730).
Motif-finding analysis
The transcription factor binding specificities were determined by RankMotif++ [31] and MEME [30]. S. pombe promoter sequences 1000 bp upstream of the translational start site were used for these motif-finding algorithms. For MEME, promoter sequences of genes with various log ratio thresholds from expression microarray and ChIP-chip experiments were input into the MEME online server. RankMotif++ was applied to the entire expression microarray data since its motif-searching algorithm is threshold independent. The consensus sequences of the transcription factor binding sites were displayed by submitting the position weight matrices obtained from RankMotif++ analysis into the enoLOGOS online server [78].
Flocculation assays
Strains were grown in flasks at 30°C for the appropriate time, and 10 ml of culture was transferred to culture tubes for strains with larger floc sizes. Images were acquired immediately after vigorous shaking in glass culture tubes with a Canon G10 digital camera. For strains with mild flocculation, flocs were harder to visualize in culture tubes, and therefore, were observed in 90 mm plastic petri dishes. 10–15 ml of culture was transferred to petri dishes, followed by gentle shaking [8] on an orbital low-speed shaker (Labnet International, Woodridge, NJ) at maximum speed for one hour in room temperature. Floc images in petri dishes were captured using a SPImager (S&P Robotics Inc., Toronto, ON). Deflocculation of flocculent strains was performed by the addition of 2–20% D-(+)-galactose or 10 mM EDTA. The reflocculation of the deflocculated cells was performed by washing with water, resuspending the cells in YES or EMM medium or 100 mM CaCl2 and allowing the culture to sit for 30 min at room temperature.
For the overexpression of pfl+ genes, the strains were inoculated at a concentration of 107 cells in 100 ml of EMM without thiamine and cultured for 3 days at 30°C. For the weaker flocculent strains (pfl2+–pfl9+), 5 ml of the 3-day culture was then inoculated into 100 ml of fresh EMM without thiamine and incubated for another 3–4 days at 30°C followed by the petri dish flocculation assay as described above. Fresh EMM medium was added on the third day to prevent cells from remaining in stationary phase. Flocculation assays for the more flocculent overexpression strains were similarly carried out except the induction times were less than three days and did not require refeeding with fresh EMM medium. It should be noted that the empty vector control cells also eventually flocculate after refeeding with fresh EMM medium, but the onset of flocculation and flocs were delayed for several days and less pronounced, respectively, compared to the weakest flocculent overexpression strains. Wild-type strain and deletion mutants (mbx2Δ, gsf2Δ, cbf12Δ, adn2Δ and adn3Δ) were induced to flocculate by inoculating cells at a concentration of 108 cells in 100 ml of YEGlycEtOH medium and culturing for 5 days at 30°C followed by the petri dish flocculation assay as described above.
Agar adhesion and invasive growth assay
A patch of cells approximately 1/6 of a 90 mm petri dish was grown on YES medium for two days at 30°C and transferred as described in [29] onto a LNB plate (0.067 g/L yeast nitrogen base without amino acids (Bacto), 20 g/L glucose, 20 g/L agar, salts and vitamins as for EMM) with an underlying layer of YE + ALU (0.5% YE, 225 mg/L adenine, leucine, and uracil each) [79]. The plates were incubated at 30°C for 2 weeks before testing for cell-to-surface adhesion by washing cells off under a gentle stream of water and for invasive growth by rubbing the remaining cells off the agar with a finger under a stream of water. For strains showing resistance to rigorous washing by finger, a small section of the agar was cut out and observed under a Zeiss AxioScope A1 tetrad microscope (Zeiss, Thornwood, NY). Invasive growth was observed by the presence of elongated and branched cells remaining underneath the agar [29], [79].
Fluorescence microscopy
Images of GFP-tagged cells were acquired with a Zeiss AxioScope 2 microscope (Zeiss, Thornwood, NY) and Scion CFW Monochrome CCD Firewire Camera (Scion Corporation, Frederick, MD). Fluorescence intensity was quantitated using the open source software ImageJ (version 1.44) (National Institutes of Health). First, the background signal for each image was subtracted using the “Subtract Background” function (50 pixel rolling ball radius). Individual cells were then selected as regions of interest using the freehand or polygon selection tools. Using the “Set Measurements” function both the area and integrated density were determined for each selected cell (n ranged between 27 and 50). Corrected GFP intensity was determined for each cell and was defined as the quotient of integrated density/area in background subtracted images. The averaged integrated density/area measurements for a given number cells is presented as the mean corrected GFP intensity with standard deviation. Significant differences between means were calculated by the Student t-test. To view nuclei and cell wall material, cells were methanol-fixed and stained with DAPI (1 µg/ml) and calcofluor white (50 µg/ml), respectively.
Supporting Information
{kind=link}
Zdroje
1. GoossensK, WillaertR (2010) Flocculation protein structure and cell-cell adhesion mechanism in Saccharomyces cerevisiae. Biotechnol Lett 32 : 1571–1585.
2. SoaresEV (2010) Flocculation in Saccharomyces cerevisiae: a review. J Appl Microbiol 110 : 1–18.
3. SmukallaS, CaldaraM, PochetN, BeauvaisA, GuadagniniS, et al. (2008) FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast. Cell 135 : 726–737.
4. VerstrepenKJ, KlisFM (2006) Flocculation, adhesion and biofilm formation in yeasts. Mol Microbiol 60 : 5–15.
5. StratfordM (1989) Evidence for two mechanisms of flocculation in Saccharomyces cerevisiae. Yeast 5 Spec No S441–445.
6. GoossensKV, StassenC, StalsI, DonohueDS, DevreeseB, et al. (2011) The N-terminal domain of the Flo1 flocculation protein from Saccharomyces cerevisiae binds specifically to mannose carbohydrates. Eukaryot Cell 10 : 110–117.
7. TanakaN, AwaiA, BhuiyanMS, FujitaK, FukuiH, et al. (1999) Cell surface galactosylation is essential for nonsexual flocculation in Schizosaccharomyces pombe. J Bacteriol 181 : 1356–1359.
8. Van MuldersSE, ChristianenE, SaerensSM, DaenenL, VerbelenPJ, et al. (2009) Phenotypic diversity of Flo protein family-mediated adhesion in Saccharomyces cerevisiae. FEMS Yeast Res 9 : 178–190.
9. KobayashiO, YoshimotoH, SoneH (1999) Analysis of the genes activated by the FLO8 gene in Saccharomyces cerevisiae. Curr Genet 36 : 256–261.
10. TeunissenAW, SteensmaHY (1995) Review: the dominant flocculation genes of Saccharomyces cerevisiae constitute a new subtelomeric gene family. Yeast 11 : 1001–1013.
11. VerstrepenKJ, DerdelinckxG, VerachtertH, DelvauxFR (2003) Yeast flocculation: what brewers should know. Appl Microbiol Biotechnol 61 : 197–205.
12. GovenderP, DomingoJL, BesterMC, PretoriusIS, BauerFF (2008) Controlled expression of the dominant flocculation genes FLO1, FLO5, and FLO11 in Saccharomyces cerevisiae. Appl Environ Microbiol 74 : 6041–6052.
13. Purevdorj-GageB, OrrME, StoodleyP, SheehanKB, HymanLE (2007) The role of FLO11 in Saccharomyces cerevisiae biofilm development in a laboratory based flow-cell system. FEMS Yeast Res 7 : 372–379.
14. GuoB, StylesCA, FengQ, FinkGR (2000) A Saccharomyces gene family involved in invasive growth, cell-cell adhesion, and mating. Proc Natl Acad Sci U S A 97 : 12158–12163.
15. LoWS, DranginisAM (1998) The cell surface flocculin Flo11 is required for pseudohyphae formation and invasion by Saccharomyces cerevisiae. Mol Biol Cell 9 : 161–171.
16. BesterMC, PretoriusIS, BauerFF (2006) The regulation of Saccharomyces cerevisiae FLO gene expression and Ca2+ -dependent flocculation by Flo8p and Mss11p. Curr Genet 49 : 375–383.
17. LiuH, StylesCA, FinkGR (1996) Saccharomyces cerevisiae S288C has a mutation in FLO8, a gene required for filamentous growth. Genetics 144 : 967–978.
18. FichtnerL, SchulzeF, BrausGH (2007) Differential Flo8p-dependent regulation of FLO1 and FLO11 for cell-cell and cell-substrate adherence of Sacc. cerevisiae S288c. Mol Microbiol 66 : 1276–1289.
19. ShenH, IhaH, YaguchiS, TsurugiK (2006) The mechanism by which overexpression of Gts1p induces flocculation in a FLO8-inactive strain of the yeast Saccharomyces cerevisiae. FEMS Yeast Res 6 : 914–923.
20. MatsuzawaT, MoritaT, TanakaN, TohdaH, TakegawaK (2011) Identification of a galactose-specific flocculin essential for non-sexual flocculation and filamentous growth in Schizosaccharomyces pombe. Mol Microbiol 82 : 1531–1544.
21. MatsuzawaT, YoritsuneK, TakegawaK (2012) MADS Box Transcription Factor Mbx2/Pvg4 Regulates Invasive Growth and Flocculation by Inducing gsf2+ Expression in Fission Yeast. Eukaryot Cell 11 : 151–158.
22. PrevorovskyM, GrouslT, StanurovaJ, RynesJ, NellenW, et al. (2009) Cbf11 and Cbf12, the fission yeast CSL proteins, play opposing roles in cell adhesion and coordination of cell and nuclear division. Exp Cell Res 315 : 1533–1547.
23. LinderT, GustafssonCM (2007) Molecular phylogenetics of ascomycotal adhesins–a novel family of putative cell-surface adhesive proteins in fission yeasts. Fungal Genet Biol 45 : 485–497.
24. LinderT, RasmussenNN, SamuelsenCO, ChatzidakiE, BaraznenokV, et al. (2008) Two conserved modules of Schizosaccharomyces pombe Mediator regulate distinct cellular pathways. Nucleic Acids Res 36 : 2489–2504.
25. KangWH, ParkYH, ParkHM (2010) The LAMMER kinase homolog, Lkh1, regulates Tup transcriptional repressors through phosphorylation in Schizosaccharomyces pombe. J Biol Chem 285 : 13797–13806.
26. BeskowA, WrightAP (2006) Comparative analysis of regulatory transcription factors in Schizosaccharomyces pombe and budding yeasts. Yeast 23 : 929–935.
27. Hertz-FowlerC, PeacockCS, WoodV, AslettM, KerhornouA, et al. (2004) GeneDB: a resource for prokaryotic and eukaryotic organisms. Nucleic Acids Res 32: D339–343.
28. AndreishchevaEN, KunkelJP, GemmillTR, TrimbleRB (2004) Five genes involved in biosynthesis of the pyruvylated Galbeta1,3-epitope in Schizosaccharomyces pombe N-linked glycans. J Biol Chem 279 : 35644–35655.
29. DodgsonJ, AvulaH, HoeKL, KimDU, ParkHO, et al. (2009) Functional genomics of adhesion, invasion, and mycelial formation in Schizosaccharomyces pombe. Eukaryot Cell 8 : 1298–1306.
30. BaileyTL, WilliamsN, MislehC, LiWW (2006) MEME: discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res 34: W369–373.
31. ChenX, HughesTR, MorrisQ (2007) RankMotif++: a motif-search algorithm that accounts for relative ranks of K-mers in binding transcription factors. Bioinformatics 23: i72–79.
32. NurrishSJ, TreismanR (1995) DNA binding specificity determinants in MADS-box transcription factors. Mol Cell Biol 15 : 4076–4085.
33. PollockR, TreismanR (1991) Human SRF-related proteins: DNA-binding properties and potential regulatory targets. Genes Dev 5 : 2327–2341.
34. ToddRB, AndrianopoulosA (1997) Evolution of a fungal regulatory gene family: the Zn(II)2Cys6 binuclear cluster DNA binding motif. Fungal Genet Biol 21 : 388–405.
35. PrevorovskyM, AtkinsonSR, PtackovaM, McLeanJR, GouldK, et al. (2011) N-termini of fungal CSL transcription factors are disordered, enriched in regulatory motifs and inhibit DNA binding in fission yeast. PLoS ONE 6: e23650 doi:10.1371/journal.pone.0023650.
36. AligianniS, LacknerDH, KlierS, RusticiG, WilhelmBT, et al. (2009) The fission yeast homeodomain protein Yox1p binds to MBF and confines MBF-dependent cell-cycle transcription to G1-S via negative feedback. PLoS Genet 5: e1000626 doi:10.1371/journal.pgen.1000626.
37. HughesAL, ToddBL, EspenshadePJ (2005) SREBP pathway responds to sterols and functions as an oxygen sensor in fission yeast. Cell 120 : 831–842.
38. ChangYW, HowardSC, HermanPK (2004) The Ras/PKA signaling pathway directly targets the Srb9 protein, a component of the general RNA polymerase II transcription apparatus. Mol Cell 15 : 107–116.
39. PrevorovskyM, PutaF, FolkP (2007) Fungal CSL transcription factors. BMC Genomics 8 : 233.
40. Artavanis-TsakonasS, RandMD, LakeRJ (1999) Notch signaling: cell fate control and signal integration in development. Science 284 : 770–776.
41. Gomez-EscodaB, IvanovaT, CalvoIA, Alves-RodriguesI, HidalgoE, et al. (2011) Yox1 links MBF-dependent transcription to completion of DNA synthesis. EMBO Rep 12 : 84–89.
42. BachewichC, NantelA, WhitewayM (2005) Cell cycle arrest during S or M phase generates polarized growth via distinct signals in Candida albicans. Mol Microbiol 57 : 942–959.
43. BachewichC, WhitewayM (2005) Cyclin Cln3p links G1 progression to hyphal and pseudohyphal development in Candida albicans. Eukaryot Cell 4 : 95–102.
44. BensenES, Clemente-BlancoA, FinleyKR, Correa-BordesJ, BermanJ (2005) The mitotic cyclins Clb2p and Clb4p affect morphogenesis in Candida albicans. Mol Biol Cell 16 : 3387–3400.
45. Chapa y LazoB, BatesS, SudberyP (2005) The G1 cyclin Cln3 regulates morphogenesis in Candida albicans. Eukaryot Cell 4 : 90–94.
46. ChouH, GloryA, BachewichC (2011) Orthologues of the anaphase-promoting complex/cyclosome coactivators Cdc20p and Cdh1p are important for mitotic progression and morphogenesis in Candida albicans. Eukaryot Cell 10 : 696–709.
47. HusseinB, HuangH, GloryA, OsmaniA, KaminskyjS, et al. (2011) G1/S transcription factor orthologues Swi4p and Swi6p are important but not essential for cell proliferation and influence hyphal development in the fungal pathogen Candida albicans. Eukaryot Cell 10 : 384–397.
48. JinR, DobryCJ, McCownPJ, KumarA (2008) Large-scale analysis of yeast filamentous growth by systematic gene disruption and overexpression. Mol Biol Cell 19 : 284–296.
49. OfirA, HofmannK, WeindlingE, GildorT, BarkerKS, et al. (2012) Role of a Candida albicans Nrm1/Whi5 homologue in cell cycle gene expression and DNA replication stress response. Mol Microbiol 84 : 778–794.
50. HortonJD, GoldsteinJL, BrownMS (2002) SREBPs: activators of the complete program of cholesterol and fatty acid synthesis in the liver. J Clin Invest 109 : 1125–1131.
51. StraussCJ, van WykPWJ, LodoloEJ, BotesPJ, PohlCH, et al. (2007) Mitochondrial Associated Yeast Flocculation -The Effect of Acetylsalicylic Acid. Journal of the Institute of Brewing 113 : 42–47.
52. IungAR, CoulonJ, KissF, EkomeJN, VallnerJ, et al. (1999) Mitochondrial function in cell wall glycoprotein synthesis in Saccharomyces cerevisiae NCYC 625 (Wild type) and [rho(0)] mutants. Appl Environ Microbiol 65 : 5398–5402.
53. DekkerN, SpeijerD, GrunCH, van den BergM, de HaanA, et al. (2004) Role of the alpha-glucanase Agn1p in fission-yeast cell separation. Mol Biol Cell 15 : 3903–3914.
54. DekkerN, van RijsselJ, DistelB, HochstenbachF (2007) Role of the alpha-glucanase Agn2p in ascus-wall endolysis following sporulation in fission yeast. Yeast 24 : 279–288.
55. OmiK, SonodaH, NagataK, SugitaK (1999) Cloning and characterization of psu1(+), a new essential fission yeast gene involved in cell wall synthesis. Biochem Biophys Res Commun 262 : 368–374.
56. de Medina-RedondoM, Arnaiz-PitaY, ClavaudC, FontaineT, del ReyF, et al. (2010) beta(1,3)-glucanosyl-transferase activity is essential for cell wall integrity and viability of Schizosaccharomyces pombe. PLoS ONE 5: e14046 doi:10.1371/journal.pone.0014046.
57. StratfordM, CarterAT (1993) Yeast flocculation: lectin synthesis and activation. Yeast 9 : 371–378.
58. ZhangN, GardnerDC, OliverSG, StatevaLI (1999) Down-regulation of the expression of PKC1 and SRB1/PSA1/VIG9, two genes involved in cell wall integrity in Saccharomyces cerevisiae, causes flocculation. Microbiology 145(Pt 2): 309–316.
59. ImazuH, SakuraiH (2005) Saccharomyces cerevisiae heat shock transcription factor regulates cell wall remodeling in response to heat shock. Eukaryot Cell 4 : 1050–1056.
60. SopkoR, HuangD, PrestonN, ChuaG, PappB, et al. (2006) Mapping pathways and phenotypes by systematic gene overexpression. Mol Cell 21 : 319–330.
61. Alonso-NunezML, AnH, Martin-CuadradoAB, MehtaS, PetitC, et al. (2005) Ace2p controls the expression of genes required for cell separation in Schizosaccharomyces pombe. Mol Biol Cell 16 : 2003–2017.
62. DekkerN, de HaanA, HochstenbachF (2006) Transcription regulation of the alpha-glucanase gene agn1 by cell separation transcription factor Ace2p in fission yeast. FEBS Lett 580 : 3099–3106.
63. PetitCS, MehtaS, RobertsRH, GouldKL (2005) Ace2p contributes to fission yeast septin ring assembly by regulating mid2+ expression. J Cell Sci 118 : 5731–5742.
64. RusticiG, MataJ, KivinenK, LioP, PenkettCJ, et al. (2004) Periodic gene expression program of the fission yeast cell cycle. Nat Genet 36 : 809–817.
65. ConlanRS, TzamariasD (2001) Sfl1 functions via the co-repressor Ssn6-Tup1 and the cAMP-dependent protein kinase Tpk2. J Mol Biol 309 : 1007–1015.
66. SongW, CarlsonM (1998) Srb/mediator proteins interact functionally and physically with transcriptional repressor Sfl1. EMBO J 17 : 5757–5765.
67. AlonU (2007) Network motifs: theory and experimental approaches. Nat Rev Genet 8 : 450–461.
68. MorenoS, KlarA, NurseP (1991) Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol 194 : 795–823.
69. JankeC, MagieraMM, RathfelderN, TaxisC, ReberS, et al. (2004) A versatile toolbox for PCR-based tagging of yeast genes: new fluorescent proteins, more markers and promoter substitution cassettes. Yeast 21 : 947–962.
70. SheffMA, ThornKS (2004) Optimized cassettes for fluorescent protein tagging in Saccharomyces cerevisiae. Yeast 21 : 661–670.
71. ForsburgSL, ShermanDA (1997) General purpose tagging vectors for fission yeast. Gene 191 : 191–195.
72. ChuaG (2009) Identification of transcription factor targets by phenotypic activation and microarray expression profiling in yeast. Methods Mol Biol 548 : 19–35.
73. SmythGK, SpeedT (2003) Normalization of cDNA microarray data. Methods 31 : 265–273.
74. SmythGK (2004) Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol 3: Article3.
75. EisenMB, SpellmanPT, BrownPO, BotsteinD (1998) Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci U S A 95 : 14863–14868.
76. SaldanhaAJ (2004) Java Treeview–extensible visualization of microarray data. Bioinformatics 20 : 3246–3248.
77. BuckMJ, NobelAB, LiebJD (2005) ChIPOTle: a user-friendly tool for the analysis of ChIP-chip data. Genome Biol 6: R97.
78. WorkmanCT, YinY, CorcoranDL, IdekerT, StormoGD, et al. (2005) enoLOGOS: a versatile web tool for energy normalized sequence logos. Nucleic Acids Res 33: W389–392.
79. Amoah-BuahinE, BoneN, ArmstrongJ (2005) Hyphal Growth in the Fission Yeast Schizosaccharomyces pombe. Eukaryot Cell 4 : 1287–1297.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 12
Nejčtenější v tomto čísle
- Population Genomics of Sub-Saharan : African Diversity and Non-African Admixture
- Dnmt3a Protects Active Chromosome Domains against Cancer-Associated Hypomethylation
- Excessive Astrocyte-Derived Neurotrophin-3 Contributes to the Abnormal Neuronal Dendritic Development in a Mouse Model of Fragile X Syndrome
- Pre-Disposition and Epigenetics Govern Variation in Bacterial Survival upon Stress