
FliZ Is a Global Regulatory Protein Affecting the Expression of Flagellar and Virulence Genes in Individual Bacterial Cells
Heterogeneity in the expression of various bacterial genes has been shown to result in the presence of individuals with different phenotypes within clonal bacterial populations. The genes specifying motility and flagellar functions are coordinately regulated and form a complex regulon, the flagellar regulon. Complex interplay has recently been demonstrated in the regulation of flagellar and virulence gene expression in many bacterial pathogens. We show here that FliZ, a DNA-binding protein, plays a key role in the insect pathogen, Xenorhabdus nematophila, affecting not only hemolysin production and virulence in insects, but efficient swimming motility. RNA-Seq analysis identified FliZ as a global regulatory protein controlling the expression of 278 Xenorhabdus genes either directly or indirectly. FliZ is required for the efficient expression of all flagellar genes, probably through its positive feedback loop, which controls expression of the flhDC operon, the master regulator of the flagellar circuit. FliZ also up - or downregulates the expression of numerous genes encoding non-flagellar proteins potentially involved in key steps of the Xenorhabdus lifecycle. Single-cell analysis revealed the bimodal expression of six identified markers of the FliZ regulon during exponential growth of the bacterial population. In addition, a combination of fluorescence-activated cell sorting and RT-qPCR quantification showed that this bimodality generated a mixed population of cells either expressing (“ON state”) or not expressing (“OFF state”) FliZ-dependent genes. Moreover, studies of a bacterial population exposed to a graded series of FliZ concentrations showed that FliZ functioned as a rheostat, controlling the rate of transition between the “OFF” and “ON” states in individuals. FliZ thus plays a key role in cell fate decisions, by transiently creating individuals with different potentials for motility and host interactions.
Published in the journal:
. PLoS Genet 9(10): e32767. doi:10.1371/journal.pgen.1003915
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1003915
Summary
Heterogeneity in the expression of various bacterial genes has been shown to result in the presence of individuals with different phenotypes within clonal bacterial populations. The genes specifying motility and flagellar functions are coordinately regulated and form a complex regulon, the flagellar regulon. Complex interplay has recently been demonstrated in the regulation of flagellar and virulence gene expression in many bacterial pathogens. We show here that FliZ, a DNA-binding protein, plays a key role in the insect pathogen, Xenorhabdus nematophila, affecting not only hemolysin production and virulence in insects, but efficient swimming motility. RNA-Seq analysis identified FliZ as a global regulatory protein controlling the expression of 278 Xenorhabdus genes either directly or indirectly. FliZ is required for the efficient expression of all flagellar genes, probably through its positive feedback loop, which controls expression of the flhDC operon, the master regulator of the flagellar circuit. FliZ also up - or downregulates the expression of numerous genes encoding non-flagellar proteins potentially involved in key steps of the Xenorhabdus lifecycle. Single-cell analysis revealed the bimodal expression of six identified markers of the FliZ regulon during exponential growth of the bacterial population. In addition, a combination of fluorescence-activated cell sorting and RT-qPCR quantification showed that this bimodality generated a mixed population of cells either expressing (“ON state”) or not expressing (“OFF state”) FliZ-dependent genes. Moreover, studies of a bacterial population exposed to a graded series of FliZ concentrations showed that FliZ functioned as a rheostat, controlling the rate of transition between the “OFF” and “ON” states in individuals. FliZ thus plays a key role in cell fate decisions, by transiently creating individuals with different potentials for motility and host interactions.
Introduction
Flagella are complex surface structures that serve as the primary means of locomotion in many bacterial species and allow many bacterial pathogens to adhere to and invade cells and in some cases to secrete virulence factors [1]. More than 50 genes are involved in the biogenesis and function of a flagellum in Escherichia coli or Salmonella enterica serovar Typhimurium (S. Typhimurium) [2]. Flagellar gene expression is sequential mirroring the timing of the assembly process [2], [3]. First expressed is the class I operon, flhDC, the products of which, FlhD4C2 heterohexamers, are required for the expression of all other flagellar genes [4]–[6]. The E. coli FlhD4C2 complex activates class II operons, including the structural genes for flagellar hook-basal body components (a type III secretion system) and the alternative sigma factor FliA [7]. The fliA gene is the first gene of the fliAZY operon in E. coli and S. Typhimurium and its product, sigma 28, directs transcription of the class III genes encoding the filament protein called flagellin, hook-associated proteins, motor proteins and various chemotaxis proteins [8]. The central channel of the flagellar apparatus is thought to serve as a passage for both flagellar component proteins and for the flagellar regulatory protein FlgM, an anti-sigma-28 factor [5], [9]. Thus, the accumulation of FlgM in the cell due to the prevention of its export blocks the transcription of class III genes, including that encoding flagellin. Two other genes within the flagellar regulon, fliT and fliZ, have been shown to regulate class II gene transcription in S. Typhimurium [10]. Disruption of the fliT gene increases class II gene transcription, whereas disruption of the fliZ gene decreases class II gene transcription, with no effect on class I transcription, suggesting that FliT and FliZ are negative and positive regulators, respectively. The type III secretion chaperone FliT has been shown to act as an anti-FlhD4C2 factor preventing the formation of the FlhD4C2-DNA complex and inhibiting its binding to class II promoters [11].
FliZ is encoded by a gene in the fliA operon, and orthologs are found only in the flagellar regulon of members of the family Enterobacteriaceae. The precise mechanism of action of FliZ remains unclear. It has been reported to activate class II flagellar gene expression and there is some evidence that it is involved in the posttranslational regulation of FlhD4C2 activity in S. Typhimurium [12]. However, the FliZ protein contains a region resembling the core DNA-binding domain of phage integrases [13], [14], suggesting that it may play a direct role in the regulation of transcription. Indeed, a primary study in Xenorhabdus nematophila (Enterobacteriaceae) showed that FliZ activated the transcription of class II flagellar genes by direct binding to the flhDC promoter [13]. Another study showed that FliZ indirectly activated flagellar gene expression in S. Typhimurium by binding directly to nlpC promoter and repressing the transcription of the associated ydiV gene, which encodes an anti-FlhDC factor [15].
Flagellar regulators, such as FliZ, have been implicated in processes other than flagellum synthesis. FliZ has been shown to be an abundant DNA-binding protein that inhibits gene expression mediated by RpoS in E. coli by recognizing operator sequences resembling the −10 region of RpoS-dependent promoters [14]. Previous studies have also shown that fliZ mutation significantly reduces hilA expression and intestinal S. Typhimurium colonization in mice [16]. Indeed, indirect regulation by FliZ has been shown to upregulate the expression of the SPI1 type three secretion system (T3SS) in S. Typhimurium, where FliZ controls HilD protein activity upstream from the HilC/RtsA/HilA transcriptional cascade [17].
As in S. Typhimurium, FliZ has been shown to mediate the coordinate regulation of flagellum synthesis and virulence in the insect pathogen X. nematophila [13]. Xenorhabdus nematophila displays complex interactions with invertebrates, has a symbiotic lifestyle with nematodes of the genus Steinernema and is pathogenic to insect larvae [18], [19]. The successful colonization of two invertebrate hosts requires Xenorhabdus to cope with shifting host environments, by interconnecting the various gene networks [19]. Various regulatory proteins of Xenorhabdus are involved in host interactions, but it has been shown that the fliAZ operon plays a central role in controlling lipase and hemolysin production and in motility and full virulence in insects [13]. Indeed, FliA coordinates the expression of class III flagellar genes, such as the flagellin-encoding gene fliC, and the two non-flagellar genes, xlpA and xrtA, encoding a lipase and a protease, respectively [20], whereas FliZ binds directly to the promoter regions of two different hemolysin-encoding operons, xaxAB [21] and xhlBA [22], activating the transcription even in the absence of the FlhD4C2 complex [13]. A real-time analysis of virulence gene expression during insect infection revealed that the expression of FliZ-dependent hemolysin genes coincided with the increase in iron availability detected at the time of insect death, suggesting that iron availability is a signal governing the adaptation of X. nematophila to changes in host environments. Interestingly, this study also revealed that the expression of the fliC and xaxAB genes in Xenorhabdus was heterogeneous at the individual cell level [23].
In 1976, Spudich and Koshland reported the existence of “non-genetic individuality” in monitoring the swimming behavior of S. Typhimurium at the level of individual cells [24]. More recently, the molecular origin of the temporal variations of chemotaxis system signaling between individual bacteria was reinvestigated, resulting in the demonstration of a role for the relative concentration of a key chemotaxis network component, CheR [25]. Furthermore, high levels of stochastic phenotypic variation have been reported for flagellar genes in S. Typhimurium [26]–[28] and Bacillus subtilis [29]. We show here that FliZ plays a key role in Xenorhabdus, not only in hemolysin activities and full virulence in insects, but also in efficient swimming motility. We demonstrate that efficient expression of the entire flagellar regulon requires the FliZ-dependent positive feedback loop controlling expression of the master operon flhDC. We also show that expression of the flagellin and FliZ-dependent hemolysin genes is heterogeneous, differing between individual cells, and that a FliZ threshold controls the rate of transition between the OFF and ON states of FliZ-dependent gene expression at the single-cell scale. FliZ-modulated bimodal gene expression generated a mixed population of cells, with different levels of FliZ-dependent gene expression, resulting in the transient production of individuals with different potentials in terms of host interactions.
Results
FliZ is a Global Regulatory Protein Affecting the Transcription of Genes Encoding Flagellar and Non-flagellar Proteins
Our previous transcriptional analysis revealed that FliZ was required for the coupling of motility and hemolysin expression in X. nematophila and that FliZ bound directly to the promoter regions of the hemolysin and flhDC operons, functioning as an activator [13]. We investigated the whole FliZ regulon using RNA-Seq analysis to compare the gene expression profiles of the wild type strain F1 and the isogenic fliZ mutant at mid-exponential growth phase (OD540 = 0.5). More than 73 million Illumina sequences (36-base reads) were obtained for each sample. More than 82% of these sequences were of sufficiently high quality and could be mapped to at least one site in the X. nematophila F1 genome [30]. Transcriptomes were compared for each annotated feature between the wild type strain and the fliZ mutant (GEO accession number: GSE47365). We observed significant differences in expression between the fliZ mutant and the wild type strain for 278 coding sequences (|log2 fold change| ≥1; adjusted P-values≤0.05; Table S1), 235 of which were downregulated and 43 of which were upregulated in the fliZ mutant relative to the wild type strain. These genes were either isolated or clustered into 23 genomic regions scattered throughout the bacterial chromosome. The genes on which fliZ inactivation had the strongest effect are listed in Table 1.
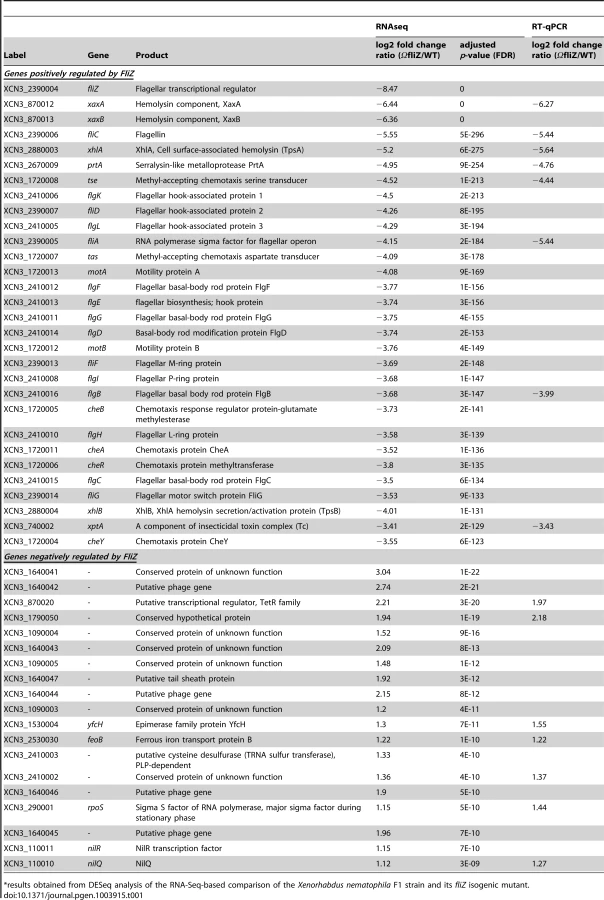
The protein-coding genes significantly downregulated in the fliZ mutant included all 47 flagellar protein-encoding genes, clustered in three flagellar regions (loci 13, 14 and 17; Table S1). Many other non-flagellar functional clusters were also upregulated by FliZ (i) the 14 genes (xcnA–N; locus 15) required for the synthesis of xenocoumacin, the major antimicrobial compound produced by X. nematophila [31]; (ii) the pax cluster (locus 10) encoding enzymes involved in synthesis of the Pax antimicrobial cyclolipopeptide [32], [33] and (iii) all 16 genes (locus 23) encoding putative components of a type VI secretion system [34], [35]. Non-flagellar genes directly regulated by FliZ, such as those encoding hemolysins (XaxAB and XhlAB) [13], were identified, as expected. However, more surprisingly, xptA1, encoding XptA1, the active component of a high-molecular weight protein toxin complex (Tc), was found to be strongly regulated by FliZ, and XptA2, a protein without insecticidal activity detected [36], was found to be weakly regulated by FliZ.
FliZ was initially described as a transcriptional activator, but we also identified individual genes and clusters of genes downregulated by this protein (Tables 1 and S1). Most of these downregulated genes are annotated as encoding hypothetical proteins of unknown function. However, FliZ also represses the transcription of a prophagic region (loci 11 and 12). The rpoS gene encoding the sigma factor σS was also found to be repressed by FliZ, consistent with the observed interference of FliZ with the expression of σS-dependent genes in E. coli [37]. We also found that FliZ downregulated the nilQR locus encoding NilR, a DNA-binding protein that, in turn, represses the transcription of nilAB and nilC required for colonization of the nematode host [38]. For validation of our differential RNA-Seq analysis, the expression level for 15 genes encoding factors potentially involved in the lifecycle of Xenorhabdus were also determined by real-time RT-PCR (Table 1). The fold-changes obtained with the two techniques were very similar, yielding a correlation coefficient (R2) of 0.96 (Table 1, Figure S1).
Taken together, these results demonstrate that FliZ is required for the efficient expression of the entire flagellar regulon in Xenorhabdus. FliZ also serves other functions, as a positive or negative regulator of the expression of numerous genes encoding non-flagellar proteins potentially involved in key steps of the Xenorhabdus lifecycle.
FliZ Inactivation Leads to a Pleiotropic Phenotype in Xenorhabdus
Mutations in the flhDC and fliAZ operons of X. nematophila have been shown to affect swimming and swarming motilities, lipase and protease production, hemolysis and insect virulence [13], [20], [39]. However, the relative impacts of these regulatory factors on phenotypes remain unclear. We tried to elucidate the role of FliZ, by comparing the phenotypic characteristics of the wild type and of flhD, fliAZ and fliZ mutants with and without fliZ expression, using an inducible Ptet-fliZ construct (Table 2). No difference was observed between strains in terms of antibiotic production, lecithin degradation and bromothymol blue adsorption. As expected [23], the fliZ mutant had no hemolytic activity on sheep blood agar and displayed an attenuated virulence phenotype in insects similar to that of the fliAZ and flhD mutants (Figure S2). However, the fliZ mutant was unable to hydrolyze Tween 20 and presented a weak ability to swim on motility agar plates; both these phenotypes are known to be FliA-mediated [20]. The ectopic expression of FliZ complemented all affected phenotypes in the fliZ mutant, but it restored only hemolytic activity in the flhD and fliAZ mutants (Table 2). The absence of complementation for lipase production and motility in these strains indicates that FliZ acts indirectly on these FliA-dependent phenotypes, through the positive feedback loop exerted by FliZ on flhD expression [13]. These data demonstrate that FliZ plays a key role, not only in hemolysin activity and full virulence in insects, but also in efficient motility and lipase activity in Xenorhabdus.

The Xenorhabdus fliZ Mutant Is Impaired for Flagellin Production
The fliZ mutants of E. coli and S. Typhimurium are fully motile and display only modest decreases in flagellar gene expression when grown in LB medium [15]. By contrast, the motility diameter developed by the Xenorhabdus fliZ mutant growing on LB motility agar plates was smaller than that obtained with the wild type strain, by a factor of 3.5 (Figure 1A and 1B). However, unlike the polar fliAZ mutation, fliZ deletion did not fully abolish the swimming capacity of Xenorhabdus. The fliZ (Ptet-fliZ) strain recovered full motility in the presence of the inducer anhydrotetracycline (aTc) (Figure 1A and 1B). We then investigated whether the motility defect of the fliZ mutant resulted from lower levels of flagellin production. The amount of flagellin produced by the fliZ mutant, as estimated by ELISA, was smaller than that produced by the wild type, by a factor of 3.2, and was similar to the amount of FliC detected in the non motile fliAZ strain (Figure 1C). As expected, expression of the Ptet-fliZ construct in the fliZ mutant fully restored flagellin production. We therefore used a PfliC-gfp[AAV] construct as a reporter for the level of expression of the flagellin gene. The GFP fluorescence signal was recorded in a bulk assay, on wild type, fliAZ and fliZ mutant cells grown for 15 h. As expected, GFP fluorescence was undetectable in the fliAZ mutant and reduced in the fliZ mutant by a factor 15 when compared to wild type (Figure 1D). Complementation assays, performed with a plasmid carrying both PfliC-gfp[AAV] and Ptet-fliZ constructs, also resulted in high levels of GFP fluorescence if the fliZ strain was cultured in the presence of the aTc inducer. Together with the results of RNA-Seq experiments, these data demonstrate that the motility defect of the fliZ mutant results from a global decrease in the expression of flagellar genes, resulting in the production of low amounts of the flagellin monomer.

The Expression of FliZ-Dependent Hemolysin and Class II–III Flagellar Genes Is Heterogeneous at the Single-Cell Level
Microscopic observations performed during insect infection have revealed that the expression of the fliC and xaxAB genes in Xenorhabdus is heterogeneous differing between individual cells [23]. We investigated the heterogeneity of fliC, xaxAB and xhlBA gene expression during bacterial growth in vitro, by measuring the expression of these genes in the wild type strain carrying the PfliC-gfp[AAV], PxaxAB-gfp[AAV] or PxhlBA-gfp[AAV] fusion by flow cytometry. Two distinct populations (OFF [GFP-negative cells] and ON [GFP-positive cells]) were visualized, providing evidence of a bimodal distribution of cells, in terms of the expression of flagellin and hemolysin genes, in Xenorhabdus (Figure 2 and S3).

The fliZ mutant of Xenorhabdus is less motile and has lower levels of flagellar protein than the wild type. It is therefore possible that the motility phenotype is supported by the expression of the flagellar regulon by only a small number of cells. We tested this hypothesis, by measuring the expression of the PfliC-gfp[AAV] fusion gene in the fliZ and fliAZ mutant strains, at single-cell resolution. We detected no fluorescent bacteria for the fliAZ mutant or the wild type strain carrying a promoter-less gfp gene used as a negative control (Figure 2). By contrast, GFP-positive cells were clearly detected for the fliZ mutant carrying PfliC-gfp[AAV], albeit in much smaller numbers than for the wild type strain, corresponding to only 0.4% of all bacteria (Figure 2). These data suggest that the low levels of fliC gene expression observed in the fliZ mutant at the whole population level (Figure 1) result from a substantial decrease in the number of bacteria expressing the flagellin gene.
In S. Typhimurium, FliZ induces a switch in the kinetics of class II flagellar gene expression [28]. We investigated whether flagellar genes from classes I, II and III were expressed with a bimodal distribution in Xenorhabdus, by monitoring the expression of flhD (class I), flgB and fliL (class II) and fliC (class III) during time-course study of the growth of the wild type strain. As for fliC, the expression of the class II genes, flgB and fliL, was bimodal throughout the growth of the bacteria (Figure 3). The percentages of bacteria corresponding to the OFF and ON populations for the fliL and fliC genes fluctuated strongly over time (Figure 3), with the ON population accounting for about 90% of all bacteria when the culture reached the early stationary phase. By contrast, only a single ON population was observed for the master operon flhDC, regardless of the growth stage considered (Figure 3). Thus, heterogeneous gene expression is intrinsic to the flagellar cascade and not due to external factors affecting flhDC expression.

We evaluated the role of the FliZ positive feedback loop in the observed bimodal pattern of expression, by quantifying flagellar gene expression in the fliZ mutant, in which the FliZ feedback loop is inactivated. As previously observed, the percentage of fliC-expressing fliZ mutant bacteria did not exceed 1%, regardless of the growth stage considered. By contrast, the class II genes, flgB and fliL, were expressed with a bimodal distribution in both the wild type and fliZ mutant strains. However, GFP-positive populations emerged later in the fliZ mutant strain. The expression of flhD remained unimodal in the fliZ mutant cells, but was much weaker (by a factor of 5) than that in the wild type strain (Figure 3). The FliZ feedback loop is, therefore, necessary for the early dynamics of class II flagellar gene expression, but dispensable for the heterogeneous expression of flagellar genes in X. nematophila.
Intracellular FliZ Levels in Xenorhabdus Determine FliZ-Dependent Gene Activation at the Single-Cell Scale
The FliZ feedback loop is not directly involved in the generation of bimodal expression patterns for class II flagellar genes, but FliZ may exert its activity through positive control over flagellar gene expression, mediating the transition between OFF and ON populations. We tested this hypothesis, by carrying out complementation assays with the plasmid carrying both PfliC-gfp[AAV] and Ptet-fliZ constructs. The addition of aTc led to a dose-dependent increase in the proportion of cells belonging to the ON population, which reached more than 96% for the highest concentrations of the inducer (Figure 4). However, the pattern of gene expression remains bimodal overtime and more specifically for the intermediate concentration of aTc (5 to 10 ng/ml). A slight increase in the amount of aTc, from 2.5 to 5 ng/ml, induced a shift of one third of the cells into the ON state. These results clearly demonstrate rheostatic control, by FliZ, of the rate of transition between OFF and ON states of flagellin gene expression at the individual scale. In contrast, the complementation assays of the flhD mutant with the plasmid carrying both Ptet-flhDC and PfliC-gfp[AAV] causes a more homogeneous response in cell population where almost all cells are either ON or OFF (Figure S4).

These findings also suggest that the FliZ-modulated dynamic heterogeneity in flagellar gene expression may give rise to an “OFF” state in which the FliZ regulon is switched off, and an “ON” state in which FliZ-dependent genes are expressed in a controlled manner. For the validation of this hypothesis, we separately quantified the levels of FliZ-dependent gene transcripts in ON and OFF populations, after cell sorting. The wild type strain carrying the PflgB-gfp[AAV] fusion displayed a constant bimodal pattern of expression, regardless of the growth phase considered (see Figure 3). We therefore used this strain to separate GFP-negative and GFP-positive bacterial cells by fluorescence-activated cell sorting (Figure 5A). As expected, the sorting of bacteria in the early exponential growth phase (EEP) showed flgB transcript levels to be eight times higher in the GFP-expressing population than in the GFP-negative population. A slight decrease in flhD transcription was found to be associated with significant decreases in the levels of expression of all the class II and class III flagellar genes studied, by factors of 4 to 80 (for fliC) in non fluorescent cells with respect to GFP-expressing cells. Non-flagellar genes encoding hemolysins (XaxAB and XhlBA) or protease PrtA/XrtA, which were previously shown to be upregulated by FliZ in our RNA-Seq analysis (Table 1), also displayed significant downregulation in GFP-negative bacterial cells. Only one gene downregulated by FliZ, feoB, displayed significantly higher levels of expression in non fluorescent than in fluorescent cells. However, the seven FliZ-repressed genes, including the genes encoding the regulators RpoS and NilQ, were only slightly more expressed in the OFF state (Figure 5B). Overall, these data indicate that the two cellular states resulting from bimodal expression of the flagellar cascade express the FliZ regulon differentially. They also suggest that the amount of FliZ present within the cell governs its fate.

Discussion
Despite increasing numbers of studies, we still know little about the precise role of FliZ in the regulation of flagellar gene expression and the coupling of flagellar regulation with the expression of virulence factors in pathogenic bacteria. This study provides comprehensive insight into the FliZ regulation circuit in individual cells of Xenorhabdus nematophila. RNA-Seq analysis revealed that FliZ was required for the efficient expression of all flagellar genes, through the positive feedback loop controlling the expression of flhDC, the master regulator of the flagellar cascade. FliZ was also found to up - or downregulate the expression of many genes encoding non-flagellar proteins potentially involved in key steps of the Xenorhabdus lifecycle (see the proposed model in Figure 6A). As already observed in the course of insect infection [23], we demonstrate here that the FliZ-dependent regulon, with the exception of the flhDC operon, is expressed in a bimodal manner in exponentially growing bacteria, leading to the establishment of subsets of cells in which FliZ-dependent genes are expressed at high or low levels (see Figure 6B). According to our model, FliZ controls the rate of transition between the “OFF” and “ON” states at the individual scale.

FliZ Is Required for the Efficient Expression of All Flagellar Genes and for Full Motility
X. nematophila FliZ is a transcriptional regulator that binds in vivo to unspecified regions of DNA upstream from the flhDC master operon, thereby exerting positive feedback on fliAZ expression [13]. The RNA-Seq analysis described here demonstrated a positive impact of FliZ on the expression of all the genes belonging to the flagellar cascade. Gradual decreases in the expression levels of class I, II and III flagellar genes were observed in the fliZ mutant (Table S1). It is likely that the lower level of FlhD in absence of FliZ feedback decreases class II gene expression that in turn prevents late class III promoter transcription through the action of FlgM control (see legend of Figure 6B for more details). Here, we also showed that the effects of the FliZ positive feedback loop on flhDC expression are critical for efficient motility in Xenorhabdus. Moreover, ectopic fliA expression in the fliZ mutant did not restore the full motility of X. nematophila (G. Jubelin, unpublished data), highlighting the key role of the FliZ protein in flagellum-driven motility.
As mentioned above, there are several major differences between the flagellar regulation circuit of X. nematophila and those of E. coli and S. Typhimurium [13]. One of these differences concerns the regulatory elements in the fliAZ promoter region. Unlike the fliAZ operon in Xenorhabdus, which is controlled principally by FlhDC [13], the fliAZ operons of E. coli and S. Typhimurium have two promoters: a class II promoter, which is recognized by the sigma 70 RNA polymerase in the presence of the FlhD4C2 activator complex, and a class III promoter, which is recognized by the FliA/sigma 28 RNA polymerase [7], [40]. However, the role of the class III promoter remains a matter of debate, because a recent study showed that the class III S. Typhimurium fliA transcript was not significantly translated when FliZ was produced from both the class II and class III transcripts [41]. The role of FliZ in the regulation of the flagellar circuit also differs considerably between the various enterobacterial species studied. Indeed, FliZ upregulates motility in S. Typhimurium [15], whereas it slightly represses motility in E. coli [37]. Like that of X. nematophila, the FliZ of E. coli interferes with flhDC expression by binding to a sequence downstream from the transcriptional start site, which resembles the −10 element of a cryptic σS-dependent promoter [37]. However, this consensus sequence is not found in the promoter region of the X. nematophila flhD gene. In S. Typhimurium, FliZ activates flagellar gene expression indirectly, by binding to the promoter region of the nlpC operon, repressing the transcription of this operon, which also controls expression of the ydiV gene encoding an anti-FlhDC factor active in minimal media [15]. However, no ydiV homologs are present in the X. nematophila genome. FliZ may exert direct or indirect feedback on flhDC expression in many bacterial species, but its overall impact on motility behavior and the expression of FliZ-targeted genes differs considerably between motile bacterial species. This led us to examine the primary function of FliZ in the flagellar cascade network more closely and to suggest that changes to the FliZ regulation circuit may play a key role in evolution, allowing motile bacterial species to adapt to their specific ecological niches.
FliZ-Modulated Bimodality and Control of Flagellar Gene Expression
Single-cell analysis revealed unexpected heterogeneity in gene expression in clonal bacterial populations in which particular gene circuits were either ON or OFF in individuals. Population heterogeneity has been reported for various phenotypes, including competence for sporulation, DNA uptake, biofilm formation and persistence in the presence of antibiotic treatment (see [42] for a review). Heterogeneity has also been reported for B. subtilis, in which a motile state and a sessile state coexist in growing bacterial populations [29]. Moreover, the use of tools favoring enrichment in noisy promoters in S. Typhimurium has revealed that two promoter sequences regulating genes involved in flagellum synthesis, fliC and flgK, display the highest levels of variation [27]. Noise can be exploited under certain conditions, to generate phenotypic heterogeneity. In the presence of positive regulatory feedback, a graded expression can be converted to a binary response, in which cells express a certain gene at high or low levels [42]. At the population level, this switch-like behavior may result in a bimodal distribution of gene expression, with the stable propagation of these differences to daughter cells. This type of gene expression pattern is commonly referred to as bistability [42]. We found that the expression patterns of class II and III flagellar genes were bimodal in X. nematophila. However the phenomenon we observed in X. nematophila is not a true bistable mechanism as the bimodality is transient for the expression of some flagellar genes (Figure 3). Surprisingly, single-cell analysis revealed differences in the expression dynamics of two class II genes, both of which were strictly FlhD-dependent. Expression of the flgB-L operon remained bimodal over time, whereas the fliL-Q promoter was switched on in almost all cells in early stationary phase (Figure 3). As the transition times are probably random, noisy expression at the population level probably results in cells expressing class II genes at different times. By contrast, the expression pattern of the master operon flhDC, which is controlled by numerous regulators [19], remained unimodal over time. Model-based predictions suggest that ‘democratic’ networks, in which a large number of genes mutually regulate each other, might limit variation in information flow by facilitating the emergence of a consensus decision. By contrast, ‘autocratic’ subnetworks may permit variation in information flow, by allowing expression levels within key regulatory hubs to differ between individuals in a population, resulting in different phenotypes [43]. Because the single class I operon, comprising the flhDC genes, is the master operon of the flagellar transcriptional hierarchy, these predictions may explain the noisy expression of the class II flagellar genes.
In the absence of X. nematophila FliZ, class II gene expression is induced later than in wild type cells and follows a bimodal pattern (Figure 3). These results also show that FliZ does not primarily induce heterogeneity in the expression of the class II flagellar genes in X. nematophila. A similar transient heterogeneity in class II flagellar gene expression has been reported for wild type cells of S. Typhimurium. By contrast to our observations in Xenorhabdus, fliZ mutation in S. Typhimurium causes a homogeneous response in the individual cells of the population [28]. The authors of this previous study suggested that the autogeneous FliA positive feedback loop resulted in heterogeneous expression from class II and III promoters suggesting an important role of FliZ in regulating flagellum assembly. As X. nematophila FliA has no effect on transcription of the fliAZ operon [13], this model cannot be applied to X. nematophila. Instead, we propose a model for X. nematophila in which the amount of FliZ within the cell has a major effect, fine-tuning the dynamics of the bimodality of flagellar gene expression. This model is based on the response of a cell population to a gradient of FliZ and shows that FliZ exerts rheostatic control over the rate of transition between the OFF and ON states of flagellin production (Figure 4). In the presence of FliZ-mediated positive feedback (Figure 6B), the noisy expression of class II flagellar genes is rapidly converted to a transient binary response, in which most of the cells strongly express class II and III flagellar genes, resulting in a motility phenotype at the population level. In the fliZ mutant, the weaker input of FlhD delays class II gene expression, probably resulting in a smaller number of cells concomitantly expressing class II flagellar genes. The heterogeneity of cellular dynamics and lower output would be expected to impair completion of the basal body-hook structure, leading in turn to the downregulation of class III gene expression through FlgM, resulting in a smaller number of motile cells (Figure 6B). This might explain why about 50% of the cells expressed class II genes whereas less than 1% of the cells of the fliZ mutant expressed the fliC gene. This scenario would account for the weak flagellum-driven motility of the fliZ mutant, due to the expression of class III genes by a much smaller proportion of the bacteria in the population.
Impact of FliZ-Modulated Bimodality on Bacterial Phenotypes and Interactions with the Host
In addition to its role in flagellar regulation, FliZ has been shown to regulate the expression of a number of non-flagellar genes, either directly or indirectly, in X. nematophila [13] and other enteric bacteria. These genes include the pathogenicity island 1 (SPI1) genes [16], [44], [45] and type 1 fimbrial genes [46] in S. Typhimurium. Our RNA-Seq analysis revealed that FliZ up - or downregulated the expression of genes encoding many non-flagellar proteins potentially involved in key steps of the Xenorhabdus lifecycle. As expected, FliZ was found to upregulate hemolysin gene expression by binding directly to the xaxAB and xhlAB promoter regions [13]. Through positive feedback on fliAZ expression, FliZ also modulates the expression of the FliA-dependent gene prtA (also called xrtA), which encodes a protease [20], [47]. The expression of the FliA-dependent lipase gene, xlpA [20], [48], was not FliZ-dependent in our RNA-Seq analysis of exponentially growing bacteria, but significant differences were observed in a stationary phase assay (A. Lanois, unpublished data). We have yet to identify the bacterial factors potentially accounting for the delayed virulence pattern observed with the fliZ (Figure S2) and flhD mutants [39]. We previously showed that the FliZ-dependent hemolysin XaxAB, which strongly induces necrosis and apoptosis in insect immunocompetent cells [21], was not required for full virulence. One interesting candidate is XptA, a high-molecular weight toxin complex (Tc) protein with insecticidal effects found in Xenorhabdus and Photorhabdus [49]. Indeed, the Tc makes a major contribution to Xenorhabdus virulence, as demonstrated by the virulence defect of an xptD1 mutant [50]. Thus, the loss of expression of xptA in the fliZ mutant, as revealed by our RNAseq data probably explains its attenuated virulence phenotype.
The repressive effects of FliZ are weak in X. nematophila. The genes annotated as FliZ-repressed include two genes encoding regulators, RpoS, the sigma factor σS, and NilR, which is indirectly involved in the colonization of the nematode host by Xenorhabdus [38], [51]. The rpoS mutation in X. nematophila is also associated with enhanced flagellum-driven motility [38], [51]. In E. coli, FliZ plays a key role in determining cell lifestyle: the FlhDC-controlled flagellum-based motility or a σS-dependent adhesive-sedentary lifestyle [37], [52]. The transition between the motile state of Xenorhabdus in insects and its mutualistic state, in which it adheres to the intestinal region of a soil-dwelling nematode, may also be regulated by interplay between RpoS and FliZ. However, the RpoS-dependent regulon has yet to be deciphered in X. nematophila.
Finally, we showed, by a combination of fluorescence-activated cell sorting and RT-qPCR quantification, that the two subpopulations coexisting during growth in vitro display differential expression of almost all the messenger RNA markers of the FliZ regulon. In vivo real-time expression analysis with an unstable GFP monitoring system has shown that FliZ target genes are upregulated just before the death of the insect, with expression levels peaking, at population level, in the larval cadavers. In addition, microscopic observations of insect cadavers have shown that FliZ-dependent gene expression in Xenorhabdus is heterogeneous, with differences observed between individuals [23]. FliZ-modulated bimodal expression may, therefore, lead to the generation of several subtypes of cells with different virulence potentials within an isogenic population of infecting bacteria (Figure 6B). It remains unclear why Xenorhabdus generates a mixed population of cells, some of which produce flagella, whereas others do not. Bacterial flagellins elicit innate immune responses in mammals and plants [1]. However, to our knowledge, no flagellin receptor has been described in insects, and it therefore remains unclear why variation in the expression of flagellar genes might be advantageous for insect infection. In the closely related association between Photorhabdus and its nematode host, the inversion of a single promoter, mediated by a site-specific recombination event, allows the bacteria to switch from a pathogenic to a mutualistic state [53]. Further studies are therefore required, to provide a clear demonstration that the bimodal expression of virulence factors constitutes a strategy for generating specialized cell types capable of surviving in different invertebrate niches, in X. nematophila.
Materials and Methods
Bacterial Strains, Plasmids and Growth Conditions
The strains and plasmids used in this study are listed in Table S2. Bacteria were grown routinely in Luria-Bertani (LB) medium or Mot broth (1% tryptone, 0.5% NaCl, 10 mM MgSO4) at 28°C (X. nematophila) or 37°C (E. coli). For motility assays, agar plates were prepared with LB broth supplemented with 0.35% agar. Antibiotic production, bromothymol blue adsorption, lecithinase, lipolytic and hemolytic activities were assessed as previously described [54]. When required, antibiotics were used at the following final concentrations: kanamycin, 20 mg.l−1, gentamicin, 30 mg.l−1 and chloramphenicol, 20 mg.l−1 for E. coli strains and 15 mg.l−1 for X. nematophila. Ptet constructs were induced by adding anhydrotetracycline (aTc) at a final concentration of 0.2 ng.l−1, unless otherwise indicated.
Construction of the fliZ Mutant
The regions upstream and downstream from fliZ (partial fliA and putA genes, respectively) were amplified by PCR with the fliA-Xba-f and fliA-BamHI-r primers for the upstream region and the putA-BamHI-f and putA-XhoI-r primers for the downstream region. The two 700 bp fragments obtained were inserted, together with the omega interposon cassette from pHP45-ΩCm conferring resistance to chloramphenicol, into pJQ200KS, to introduce the ΩCam cassette between the two PCR fragments. The resulting plasmid, pGJ906, was then used to transform E. coli strain S17.1 and was introduced into X. nematophila F1 in a mating experiment. Cmr and Sucr exconjugants were selected on LB agar supplemented with 4% sucrose and chloramphenicol. Omega insertion was confirmed by PCR analysis and the loss of hemolytic activity of the resulting fliZ strain was checked on sheep blood agar plates.
Molecular Genetic Techniques and RNA Preparation
DNA manipulations were carried out as previously described [55]. Plasmids were introduced into E. coli by transformation and transferred to X. nematophila by conjugative mating [39]. All constructs were sequenced by Millegen (Labège, France). The primers used in this study (Eurogentec) are described in Table S3.
Total RNA was extracted with the RNeasy Protect Bacteria miniprep (for RNA-Seq experiment) or the RNeasy Micro kit (for sorted cells, from Qiagen) including DNase I incubation in accordance with the manufacturer's recommendations. For each RNA preparation, we assessed DNA contamination by carrying out a control PCR. The quantity and quality of total and messenger RNA, respectively, were assessed with a NanoDrop 2000 spectrophotometer (Thermo Scientific) and an Agilent 2100 Bioanalyzer with the RNA 6000 Nano LabChip kit (Agilent).
Material for RNA-Seq analysis was prepared by extracting total RNA from the Xenorhabdus wild type strain and the fliZ mutant grown in Mot broth (OD540 = 0.5) (six independent biological replicates per strain) and pooling equal amounts of total RNA from three replicates of the same strain together, to generate two biological samples for each strain, which were subjected to two successive rounds of ribosomal RNA depletion with the Microbe Express kit (Ambion) according to the manufacturer's instructions.
RNA Sequencing
RNA-Seq libraries were constructed with the Truseq RNA sample preparation kit from Illumina. Briefly, for each sample, 100 ng of rRNA-depleted RNA was chemically fragmented. The first cDNA strand was generated by reverse transcription with random hexamer primers and SuperScript II Reverse Transcriptase (Life Technologies) and the second strand was then synthesized. A blunt-ended double-stranded DNA was then generated by repair techniques. A single “A” nucleotide was added to the 3′ end and ligation was carried out with Illumina's indexed adapters. After 15 cycles of PCR, libraries were validated with a DNA 1000 Labchip on a Bioanalyzer (Agilent) and quantified with a KAPA qPCR kit. For each sequencing lane, two libraries were pooled in equal proportions, denatured with NaOH and diluted to 8 pM before clustering. Clustering and 50 nt single-read sequencing were performed according to the manufacturer's instructions. Image analysis and base-calling were carried out with HiSeq Control Software (HCS 1.5.15) and a RTA component (RTA 1.13.48). Finally, demultiplexing was carried out with CASAVA (1.8.1).
RNA-Seq Analysis
Transcriptomic high-throughput sequencing data were analyzed with a bioinformatic pipeline implemented within the Microscope platform [56]. The pipeline currently used is a “Master” shell script that launches the various parts of the analysis (i.e. a collection of Shell/Perl/R scripts) and checks that all tasks are completed without error. We first assessed RNA-Seq data quality by including options, such as read-trimming or the use of merging/split paired-end reads. We then mapped reads onto the contigs of the X. nematophila F1 genome sequence (accession number: CAVM000000000) with the SSAHA2 package [57], which combines the SSAHA searching algorithm (sequence information encoded in a perfect hash function) for identifying regions of high similarity, and the cross-match sequence alignment program, which aligns these regions, using a banded Smith-Waterman-Gotoh algorithm. An alignment score covering at least half of the read is required for a hit to be retained. We minimized the false positive discovery rate, by using SAMtools (v.0.1.8) to extract reliable alignments from SAM-formatted files. The number of reads matching each genomic object harbored by the reference genome was then calculated with the Bioconductor-Genomic Features package. If reads matched several genomic objects, the count number was weighted so as to keep the total number of reads constant. Finally, the Bioconductor-DESeq package [58] was used with default parameters for the analysis of raw count data and to determine whether expression levels differed between conditions. The complete dataset from this study has been deposited in the GEO database under accession no. GSE47365.
RT-qPCR Analysis
RT-qPCR was performed in two steps. First, the cDNA was synthesized from 1 µg of total RNA from each replicate used for RNA-Seq (0.2 µg of total RNA was used for sorted cells), with Super Script II Reverse Transcriptase from Invitrogen and random hexamers (100 ng.µl−1) from Applied Biosystems. We then carried out qPCR in triplicate with the LightCycler 480 SYBR Green I Master kit from Roche Diagnostics, with 1 µl of cDNA synthesis mixture (diluted 1∶100 or 1∶20) and 1 µM of specific primers for the genes studied (Table S3). The enzyme was activated by heating for 10 minutes at 95°C. All qPCRs were performed in three technical replicates, with 45 cycles of 95°C for 5 s, 60°C for 5 s and 72°C for 10 s, and were monitored with the Light Cycler 480 system (Roche). Melting curves were analyzed for each reaction and each curve contained a single peak. The recA gene was used as the reference housekeeping gene and 16S, mre or ampD was used as an internal control. The data for each sample are expressed relative to the expression level of recA, as follows [59]: . The relative expression ratio for a target gene was calculated on the basis of its real-time PCR efficiency (E) and the crossing point (CP) difference (Δ) between a sample and the control (ΔCPcontrol – sample). Crossing points (CP) for each sample and each gene were calculated with LightCycler480 (Roche) software, using second-derivative maximum analysis and the CP medians of the technical replicates for each biological sample used. All relative quantifications were assessed with REST software 2009, using the pairwise fixed randomization test with 2,000 permutations, with PCR efficiencies calculated with serial dilutions of a mixture of cDNAs [59]. This method provided a relative quantification of the expression of a target gene with respect to a reference gene, for the comparison of the wild type and fliZ mutant strains or of GFP-positive and GFP-negative cells.
Construction of Plasmids Expressing gfp[AAV] under the Control of fliC, flhD, flgB or fliL Gene Promoters and of Plasmids Expressing fliZ or flhDC under the Control of an Inducible Ptet Promoter
The construction of the PfliC-gfp[AAV], PxaxAB-gfp[AAV], PxhlBA-gfp[AAV] and PD31-gfp[AAV] fusions have been described elsewhere [23]. We used a similar method to obtain plasmids expressing the reporter gene gfp[AAV] under the control of the flhD, flgB or fliL promoter region. Briefly, DNA fragments corresponding to the flhD (576 bp), flgB (400 bp) and fliL (223 bp) promoters were amplified by PCR from F1 genomic DNA, with primers containing an EcoRI or BamHI restriction site. The PCR products were digested and inserted into the corresponding sites of pPROBE′-gfp[AAV] for flhD and pPROBE-gfp[AAV] for flgB and fliL, yielding PflhD-gfp[AAV], PflgB-gfp[AAV] and PfliL-gfp[AAV].
For Ptet-MCS construction, we amplified the PLtet o-1-MCS-tetR DNA fragment from pSS012 by PCR with the Ptet-XhoI-f and 3′PROTetSeq primers and inserted it in place of gfp[AAV] in pPROBE-gfp[AAV] digested with SalI and EcoRV. We amplified the fliZ gene by PCR from F1 genomic DNA, with the LfliZ-Eco and RfliZ-Bam primers, and inserted it into Ptet-MCS digested with EcoRI and BamHI, to generate Ptet-fliZ. The same strategy was used with flhDC operon using LflhDEco2 and RflhCBam primers to generate Ptet-flhDC. Finally, the Ptet-fliZ-tetR and Ptet-flhDC-tetR DNA fragments were amplified by PCR from Ptet-fliZ or Ptet-flhDC respectively with the Tet-fliZ-f and Tet-fliZ-r primers and inserted into PfliC-gfp[AAV] digested with SalI and SbfI, to yield the PfliC-gfp[AAV] - Ptet-fliZ construct or PfliC-gfp[AAV] - Ptet-flhDC construct.
Quantification of fliC Expression in Bacterial Populations
Wild type, fliA and fliZ strains carrying either PfliC-gfp[AAV] or PfliC-gfp[AAV] – Ptet-fliZ constructs were cultured in black-sided, clear-bottomed 96-well plates (Greiner). For each well, 20 µl of a 1/50 dilution of an overnight culture was added to 180 µl of LB supplemented with kanamycin, and 200 ng.ml−1 of aTc when required. Then, the plates were incubated, with shaking on an orbital shaker, at 28°C, in an Infinite M200 microplate reader (Tecan). Absorbance at 600 nm and GFP fluorescence intensity, with excitation at 485±4.5 nm and emission at 520±10 nm, were measured after 15 hours of growth. Specific fluorescence was obtained by dividing fluorescence units by the absorbance value. As a control, we checked that addition of aTc did not affect the expression of PfliC-gfp[AAV] fusion in the wild type strain (data not shown).
Quantification of Gene Expression in Individual Bacterial Cells by Flow Cytometry
Bacterial strains were grown in LB supplemented with kanamycin at 28°C. If necessary, aTc was added at the indicated concentrations after 3 h of growth (OD540∼0.1). At an OD540 of ∼0.5 (or OD540 = 1.8 for PxaxAB-gfp[AAV] and PxhlBA-gfp[AAV] plasmids), samples were taken, washed once with PBS, diluted and immediately analyzed by flow cytometry (FACS Canto II, BD Biosciences). For kinetic analyses, samples were taken at the indicated time points, washed once with PBS and bacteria were fixed by incubation in 2% formaldehyde in PBS for 15 minutes at room temperature. The cells were then washed once with PBS and bacterial pellets were stored at 4°C until flow cytometry analysis. Forward scatter (FSC), side scatter (SSC) and GFP parameters were set to log and bi-exponential display was used for the GFP parameter. We captured a total of 30,000 bacteria for each sample, unless otherwise indicated, and raw data were analyzed with FlowJo version 8.8.6 software (TreeStar).
Enzyme-Linked Immunosorbent Assay (ELISA)
FliC was detected by ELISA in bacterial lysates of wild type F1, fliA and fliZ strains carrying either Ptet-fliZ or the vector control Ptet-MCS. These strains were cultured in LB medium, and 200 ng.ml−1 of anhydrotetracycline (aTc, Clontech) was added, when required, during the early exponential growth phase (OD540 = 0.2). Two hours after induction, samples were taken, centrifuged and the bacterial pellets were resuspended in ultrapure water and lysed by three freeze–thaw cycles and sonication. For ELISA, microtiter plates (Maxisorp Nunc-Immuno Plate) were coated with an amount of culture supernatant (50 µl/well) equivalent to 0.1 OD540 units of bacterial lysate. The plates were incubated overnight at room temperature, washed three times with 0.05% Tween 20 in PBS (PBS-T) and blocked by incubation with 0.25% BSA in PBS-T for 2 h at room temperature. Anti-FliC antibodies [60] diluted in 1% BSA in PBS-T (1/500) were added to each well and the plates were incubated for 1 h at room temperature. The plates were washed four times with PBS-T and incubated for 1 h with peroxidase-linked donkey anti-rabbit IgG (1/5000 dilution; GE Healthcare). The plates were washed four times with PBS-T, and 100 µl of 1-Step Ultra TMB-ELISA (Pierce) solution was added to each well. Color development was stopped after 20 minutes, by adding 100 µl of 2 M H2SO4, and absorbance at 450 nm was measured with a microplate reader (Tecan Infinite 200).
Fluorescence-Activated Cell Sorting (FACS) Analysis and Subpopulation Sorting
For the sorting of GFP-negative and GFP-positive subpopulations, an overnight culture of the F1 strain containing PflgB-gfp[AAV] was washed once with PBS, diluted 1∶250 in fresh LB broth supplemented with kanamycin and incubated at 28°C for 6 h (OD540 of 0.3 to 0.8). The cells were then washed once with PBS before FACS analysis.
Flow-cytometric sorting was performed on a FACSAria II cell sorter system (Becton Dickinson). We sorted 107 GFP-negative and GFP-positive cells at 4°C, for a maximum of 2 hours. The sorting efficiency for three independent biological samples was determined by analyzing the GFP fluorescence patterns of the subpopulations obtained. Immediately after cell sorting, RNA Protect Bacteria Reagent (Qiagen) was added to the cells and RNA was extracted with the RNeasy micro kit (Qiagen) as described above and eluted in a final volume of 15 µl.
Supporting Information
{kind=link}
Zdroje
1. RamosHC, RumboM, SirardJC (2004) Bacterial flagellins: mediators of pathogenicity and host immune responses in mucosa. Trends Microbiol 12 : 509–517.
2. MacnabRM (1992) Genetics and biogenesis of bacterial flagella. Annu Rev Genet 26 : 131–158.
3. KalirS, McClureJ, PabbarajuK, SouthwardC, RonenM, et al. (2001) Ordering genes in a flagella pathway by analysis of expression kinetics from living bacteria. Science 292 : 2080–2083.
4. BartlettDH, FrantzBB, MatsumuraP (1988) Flagellar transcriptional activators FlbB and FlaI: gene sequences and 5′ consensus sequences of operons under FlbB and FlaI control. J Bacteriol 170 : 1575–1581.
5. KutsukakeK (1994) Excretion of the anti-sigma factor through a flagellar substructure couples flagellar gene expression with flagellar assembly in Salmonella typhimurium. Mol Gen Genet 243 : 605–612.
6. WangS, FlemingRT, WestbrookEM, MatsumuraP, McKayDB (2006) Structure of the Escherichia coli FlhDC complex, a prokaryotic heteromeric regulator of transcription. J Mol Biol 355 : 798–808.
7. LiuX, MatsumuraP (1994) The FlhD/FlhC complex, a transcriptional activator of the Escherichia coli flagellar class II operons. J Bacteriol 176 : 7345–7351.
8. OhnishiK, KutsukakeK, SuzukiH, IinoT (1990) Gene fliA encodes an alternative sigma factor specific for flagellar operons in Salmonella typhimurium. Mol Gen Genet 221 : 139–147.
9. HughesKT, GillenKL, SemonMJ, KarlinseyJE (1993) Sensing structural intermediates in bacterial flagellar assembly by export of a negative regulator. Science 262 : 1277–1280.
10. KutsukakeK, IkebeT, YamamotoS (1999) Two novel regulatory genes, fliT and fliZ, in the flagellar regulon of Salmonella. Genes Genet Syst 74 : 287–292.
11. YamamotoS, KutsukakeK (2006) FliT acts as an anti-FlhD2C2 factor in the transcriptional control of the flagellar regulon in Salmonella enterica serovar typhimurium. J Bacteriol 188 : 6703–6708.
12. SainiS, BrownJD, AldridgePD, RaoCV (2008) FliZ is a posttranslational activator of FlhD4C2-dependent flagellar gene expression. J Bacteriol 190 : 4979–4988.
13. LanoisA, JubelinG, GivaudanA (2008) FliZ, a flagellar regulator, is at the crossroads between motility, haemolysin expression and virulence in the insect pathogenic bacterium Xenorhabdus. Mol Microbiol 68 : 516–533.
14. PesaventoC, HenggeR (2012) The global repressor FliZ antagonizes gene expression by sigmaS-containing RNA polymerase due to overlapping DNA binding specificity. Nucleic Acids Res 40 : 4783–93.
15. WadaT, TanabeY, KutsukakeK (2011) FliZ acts as a repressor of the ydiV gene, which encodes an anti-FlhD4C2 factor of the flagellar regulon in Salmonella enterica serovar typhimurium. J Bacteriol 193 : 5191–5198.
16. LucasRL, LostrohCP, DiRussoCC, SpectorMP, WannerBL, et al. (2000) Multiple factors independently regulate hilA and invasion gene expression in Salmonella enterica serovar typhimurium. J Bacteriol 182 : 1872–1882.
17. ChubizJE, GolubevaYA, LinD, MillerLD, SlauchJM (2010) FliZ regulates expression of the Salmonella pathogenicity island 1 invasion locus by controlling HilD protein activity in Salmonella enterica serovar typhimurium. J Bacteriol 192 : 6261–6270.
18. Nielsen-LeRouxC, GaudriaultS, RamaraoN, LereclusD, GivaudanA (2012) How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr Opin Microbiol 15 : 220–231.
19. RichardsGR, Goodrich-BlairH (2009) Masters of conquest and pillage: Xenorhabdus nematophila global regulators control transitions from virulence to nutrient acquisition. Cell Microbiol 11 : 1025–1033.
20. ParkD, ForstS (2006) Co-regulation of motility, exoenzyme and antibiotic production by the EnvZ-OmpR-FlhDC-FliA pathway in Xenorhabdus nematophila. Mol Microbiol 61 : 1397–1412.
21. VigneuxF, ZumbihlR, JubelinG, RibeiroC, PoncetJ, et al. (2007) The xaxAB genes encoding a new apoptotic toxin from the insect pathogen Xenorhabdus nematophila are present in plant and human pathogens. J Biol Chem 282 : 9571–9580.
22. CowlesKN, Goodrich-BlairH (2005) Expression and activity of a Xenorhabdus nematophila haemolysin required for full virulence towards Manduca sexta insects. Cell Microbiol 7 : 209–219.
23. JubelinG, PagesS, LanoisA, BoyerMH, GaudriaultS, et al. (2011) Studies of the dynamic expression of the Xenorhabdus FliAZ regulon reveal atypical iron-dependent regulation of the flagellin and haemolysin genes during insect infection. Environ Microbiol 13 : 1271–1284.
24. SpudichJL, KoshlandDEJr (1976) Non-genetic individuality: chance in the single cell. Nature 262 : 467–471.
25. KorobkovaE, EmonetT, VilarJM, ShimizuTS, CluzelP (2004) From molecular noise to behavioural variability in a single bacterium. Nature 428 : 574–578.
26. CummingsLA, WilkersonWD, BergsbakenT, CooksonBT (2006) In vivo, fliC expression by Salmonella enterica serovar Typhimurium is heterogeneous, regulated by ClpX, and anatomically restricted. Mol Microbiol 61 : 795–809.
27. FreedNE, SilanderOK, StecherB, BohmA, HardtWD, et al. (2008) A simple screen to identify promoters conferring high levels of phenotypic noise. PLoS Genet 4: e1000307.
28. SainiS, KoiralaS, FloessE, MearsPJ, ChemlaYR, et al. (2010) FliZ induces a kinetic switch in flagellar gene expression. J Bacteriol 192 : 6477–6481.
29. KearnsDB, LosickR (2005) Cell population heterogeneity during growth of Bacillus subtilis. Genes Dev 19 : 3083–3094.
30. LanoisA, OgierJ-C, GouzyJ, LarouiC, RouyZ, et al. (2013) Draft genome sequence and annotation of the entomopathogenic bacterium, Xenorhabdus nematophila strain F1. Genome Announcements 1: e00342–00313.
31. ParkD, CiezkiK, van der HoevenR, SinghS, ReimerD, et al. (2009) Genetic analysis of xenocoumacin antibiotic production in the mutualistic bacterium Xenorhabdus nematophila. Mol Microbiol 73 : 938–949.
32. GualtieriM, AumelasA, ThalerJO (2009) Identification of a new antimicrobial lysine-rich cyclolipopeptide family from Xenorhabdus nematophila. J Antibiot (Tokyo) 62 : 295–302.
33. FuchsSW, ProschakA, JaskollaTW, KarasM, BodeHB (2011) Structure elucidation and biosynthesis of lysine-rich cyclic peptides in Xenorhabdus nematophila. Org Biomol Chem 9 : 3130–3132.
34. ChastonJM, SuenG, TuckerSL, AndersenAW, BhasinA, et al. (2011) The entomopathogenic bacterial endosymbionts Xenorhabdus and Photorhabdus: convergent lifestyles from divergent genomes. PLoS One 6: e27909.
35. OgierJC, CalteauA, ForstS, Goodrich-BlairH, RocheD, et al. (2010) Units of plasticity in bacterial genomes: new insight from the comparative genomics of two bacteria interacting with invertebrates, Photorhabdus and Xenorhabdus. BMC Genomics 11 : 568.
36. MorganJA, SergeantM, EllisD, OusleyM, JarrettP (2001) Sequence analysis of insecticidal genes from Xenorhabdus nematophilus PMFI296. Appl Environ Microbiol 67 : 2062–2069.
37. PesaventoC, HenggeR (2012) The global repressor FliZ antagonizes gene expression by sigmaS-containing RNA polymerase due to overlapping DNA binding specificity. Nucleic Acids Res 40 : 4783–4793.
38. CowlesCE, Goodrich-BlairH (2006) nilR is necessary for co-ordinate repression of Xenorhabdus nematophila mutualism genes. Mol Microbiol 62 : 760–771.
39. GivaudanA, LanoisA (2000) flhDC, the flagellar master operon of Xenorhabdus nematophilus: requirement for motility, lipolysis, extracellular hemolysis, and full virulence in insects. J Bacteriol 182 : 107–115.
40. IkebeT, IyodaS, KutsukakeK (1999) Structure and expression of the fliA operon of Salmonella typhimurium. Microbiology 145 Pt 6 1389–1396.
41. TanabeY, WadaT, OnoK, AboT, KutsukakeK (2011) The transcript from the sigma(28)-dependent promoter is translationally inert in the expression of the sigma(28)-encoding gene fliA in the fliAZ operon of Salmonella enterica serovar Typhimurium. J Bacteriol 193 : 6132–6141.
42. VeeningJW, SmitsWK, KuipersOP (2008) Bistability, epigenetics, and bet-hedging in bacteria. Annu Rev Microbiol 62 : 193–210.
43. ChalanconG, RavaraniCN, BalajiS, Martinez-AriasA, AravindL, et al. (2012) Interplay between gene expression noise and regulatory network architecture. Trends Genet 28 : 221–232.
44. GolubevaYA, SadikAY, EllermeierJR, SlauchJM (2012) Integrating global regulatory input into the Salmonella pathogenicity island 1 type III secretion system. Genetics 190 : 79–90.
45. IyodaS, KamidoiT, HiroseK, KutsukakeK, WatanabeH (2001) A flagellar gene fliZ regulates the expression of invasion genes and virulence phenotype in Salmonella enterica serovar Typhimurium. Microb Pathog 30 : 81–90.
46. SainiS, SlauchJM, AldridgePD, RaoCV (2010) Role of cross talk in regulating the dynamic expression of the flagellar Salmonella pathogenicity island 1 and type 1 fimbrial genes. J Bacteriol 192 : 5767–5777.
47. MassaoudMK, MarokhaziJ, FodorA, VenekeiI (2010) Proteolytic enzyme production by strains of the insect pathogen Xenorhabdus and characterization of an early-log-phase-secreted protease as a potential virulence factor. Appl Environ Microbiol 76 : 6901–6909.
48. RichardsGR, Goodrich-BlairH (2010) Examination of Xenorhabdus nematophila lipases in pathogenic and mutualistic host interactions reveals a role for xlpA in nematode progeny production. Appl Environ Microbiol 76 : 221–229.
49. WaterfieldNR, BowenDJ, FetherstonJD, PerryRD, ffrench-ConstantRH (2001) The tc genes of Photorhabdus: a growing family. Trends Microbiol 9 : 185–191.
50. RichardsGR, HerbertEE, ParkY, Goodrich-BlairH (2008) Xenorhabdus nematophila lrhA is necessary for motility, lipase activity, toxin expression, and virulence in Manduca sexta insects. J Bacteriol 190 : 4870–4879.
51. VivasEI, Goodrich-BlairH (2001) Xenorhabdus nematophilus as a model for host-bacterium interactions: rpoS is necessary for mutualism with nematodes. J Bacteriol 183 : 4687–4693.
52. PesaventoC, BeckerG, SommerfeldtN, PosslingA, TschowriN, et al. (2008) Inverse regulatory coordination of motility and curli-mediated adhesion in Escherichia coli. Genes Dev 22 : 2434–2446.
53. SomvanshiVS, SloupRE, CrawfordJM, MartinAR, HeidtAJ, et al. (2012) A single promoter inversion switches Photorhabdus between pathogenic and mutualistic states. Science 337 : 88–93.
54. BoemareN, ThalerJ-O, LanoisA (1997) Simple bacteriological tests for phenotypic characterization of Xenorhabdus and Photorhabdus phase variants. Symbiosis 22 : 167–175.
55. Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, et al.. (1993) Current Protocols in Molecular Biology. Sons JW, editor. New York.
56. VallenetD, BeldaE, CalteauA, CruveillerS, EngelenS, et al. (2013) MicroScope–an integrated microbial resource for the curation and comparative analysis of genomic and metabolic data. Nucleic Acids Res 41: D636–647.
57. NingZ, CoxAJ, MullikinJC (2001) SSAHA: a fast search method for large DNA databases. Genome Res 11 : 1725–1729.
58. AndersS, HuberW (2010) Differential expression analysis for sequence count data. Genome Biol 11: R106.
59. PfafflMW, HorganGW, DempfleL (2002) Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30: e36.
60. GivaudanA, BaghdiguianS, LanoisA, BoemareN (1995) Swarming and swimming changes concomitant with phase variation in Xenorhabdus nematophilus. Appl Environ Microbiol 61 : 1408–1413.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2013 Číslo 10
Nejčtenější v tomto čísle
- A GDF5 Point Mutation Strikes Twice - Causing BDA1 and SYNS2
- Dominant Mutations in Identify the Mlh1-Pms1 Endonuclease Active Site and an Exonuclease 1-Independent Mismatch Repair Pathway
- Eleven Candidate Susceptibility Genes for Common Familial Colorectal Cancer
- The Histone H3 K27 Methyltransferase KMT6 Regulates Development and Expression of Secondary Metabolite Gene Clusters