
Loss of Circulating CD4 T Cells with B Cell Helper Function during Chronic HIV Infection
The interaction between follicular T helper cells (TFH) and B cells in the lymph nodes and spleen has a major impact on the development of antigen-specific B cell responses during infection or vaccination. Recent studies described a functional equivalent of these cells among circulating CD4 T cells, referred to as peripheral TFH cells. Here, we characterize the phenotype and in vitro B cell helper activity of peripheral TFH populations, as well as the effect of HIV infection on these populations. In co-culture experiments we confirmed CXCR5+ cells from HIV-uninfected donors provide help to B cells and more specifically, we identified a CCR7highCXCR5highCCR6highPD-1high CD4 T cell population that secretes IL-21 and enhances isotype-switched immunoglobulin production. This population is significantly decreased in treatment-naïve, HIV-infected individuals and can be recovered after anti-retroviral therapy. We found impaired immunoglobulin production in co-cultures from HIV-infected individuals and found no correlation between the frequency of peripheral TFH cells and memory B cells, or with neutralization activity in untreated HIV infection in our cohort. Furthermore, we found that within the peripheral TFH population, the expression level of TFH-associated genes more closely resembles a memory, non-TFH population, as opposed to a TFH population. Overall, our data identify a heterogeneous population of circulating CD4 T cells that provides in vitro help to B cells, and challenges the origin of these cells as memory TFH cells.
Published in the journal:
. PLoS Pathog 10(1): e32767. doi:10.1371/journal.ppat.1003853
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1003853
Summary
The interaction between follicular T helper cells (TFH) and B cells in the lymph nodes and spleen has a major impact on the development of antigen-specific B cell responses during infection or vaccination. Recent studies described a functional equivalent of these cells among circulating CD4 T cells, referred to as peripheral TFH cells. Here, we characterize the phenotype and in vitro B cell helper activity of peripheral TFH populations, as well as the effect of HIV infection on these populations. In co-culture experiments we confirmed CXCR5+ cells from HIV-uninfected donors provide help to B cells and more specifically, we identified a CCR7highCXCR5highCCR6highPD-1high CD4 T cell population that secretes IL-21 and enhances isotype-switched immunoglobulin production. This population is significantly decreased in treatment-naïve, HIV-infected individuals and can be recovered after anti-retroviral therapy. We found impaired immunoglobulin production in co-cultures from HIV-infected individuals and found no correlation between the frequency of peripheral TFH cells and memory B cells, or with neutralization activity in untreated HIV infection in our cohort. Furthermore, we found that within the peripheral TFH population, the expression level of TFH-associated genes more closely resembles a memory, non-TFH population, as opposed to a TFH population. Overall, our data identify a heterogeneous population of circulating CD4 T cells that provides in vitro help to B cells, and challenges the origin of these cells as memory TFH cells.
Introduction
Follicular helper CD4 T cells (TFH) are crucial for the development of antigen-specific B cells within germinal centers (GC). TFH cells interact through co-stimulatory receptors and provide essential soluble factors (i.e. IL-4, IL-21) to promote the survival, isotype switching and selection of high affinity memory B cells [1]. Phenotypic and gene signature analysis has revealed a highly conserved molecular profile of TFH cells in humans, non-human primates (NHP) and mice, which is characterized by increased expression of Bcl-6, CXCR5, PD-1, ICOS and decreased expression of CCR7 [2]–[4]. Human TFH cells exhibit a polarized cytokine profile characterized by compromised production of TH1 cytokines and increased secretion of IL-4, IL-10 and IL-21 [5]. Although IL-21 is characterized as a “hallmark” cytokine of TFH cells, other THelper subsets produce this cytokine [6].
The origin and differentiation of TFH is unclear, as previous studies found TFH cells can derive from TH1 or TH2 cells, or independently of other CD4 lineages [7]–[9]. However, it is well established that the transcription factor Bcl-6 regulates several molecules involved in TFH development (i.e. PD-1, IL-21R, CXCR5) [10], [11]. Similarly, the fate of TFH, particularly those in the germinal center (GC-TFH), following the effector phase of the immune response is unclear. We have recently shown that NHP GC-TFH display compromised in vivo cell cycling and are prone to in vitro cell death [4]. Other studies have shown that TFH can form a memory pool found in anatomical sites outside the lymph nodes [12]. Hence, TFH cells may adopt a “central memory” phenotype or undergo cell death after the effector phase [13]. In humans, a circulating CD4 T cell population characterized by high CXCR5 expression can provide in vitro help for B cell isotype switching and shares functional characteristics with TFH cells [14]. It was proposed that these circulating cells, termed “peripheral TFH” (pTFH) could represent the memory counterparts of TFH outside the lymphoid organs. Further investigation is needed to establish a direct relationship between TFH cells and pTFH cells.
It is becoming increasingly important to understand the interplay between CD4 T cells and B cells during HIV infection, specifically with relation to the generation of broadly neutralizing antibodies. Chronic HIV/SIV infection results in profound changes in CD4 T cell dynamics in lymph nodes characterized by TFH accumulation and increased ability of non-TFH to egress the lymph node [4], [15]. How this impacts upon the dynamics of pTFH is unknown. Elucidating the biology and dynamics of pTFH, and their ability to provide B cell help may be important for our understanding of TFH memory formation during chronic infection, as well as the establishment of immune correlates reflecting the interactions between CD4 T cells and B cells within secondary lymphoid organs. This is of particular interest for monitoring clinical studies where the B cell arm of the immune system is under investigation [16].
Here we define, detect, quantify and characterize peripheral CD4 T cell populations that support B cell differentiation. We show that particular circulating CD4 T cell populations with distinct cytokine profiles have the capacity to help B cells in vitro. We further show that the frequencies of pTFH populations are significantly compromised during chronic HIV infection but can recover with antiretroviral treatment (ART), although in vitro immunoglobulin production from HIV-infected subjects both on and off ART is reduced compared to healthy subjects. Furthermore, gene expression analysis of pTFH cells and CD4 T cells in tonsil tissue suggest pTFH cells are most closely related to a non-TFH memory population within secondary lymphoid organs. Overall, our data challenge the relationship between pTFH cells and TFH memory cells.
Results
Characterization of peripheral T follicular helper (pTFH) cells
Previous studies defined a population of circulating CD4 T cells that express CXCR5, promote the differentiation of naïve B cells and induce immunoglobulin secretion in vitro [14], [17]. We further defined CXCR5high CD4 T cells from blood, analyzed their cytokine production and determined their ability to promote B cell differentiation in vitro. CXCR5high CD4 T cells were found predominantly within the CD27highCD45ROhigh CD4 T cell population (hereafter referred to as central memory (CM)), in agreement with previous studies [17]. The majority of the CXCR5high CD4 T cell population also expressed CCR7 and we found the CCR7highCXCR5high population represented 6.5+/−2.8% (mean+/−S.D.) of total CD4 T cells in healthy subjects (Figure 1A). The majority of CXCR5high cells expressed CD150. We further analyzed these cells based on expression of CCR6, which was previously used in combination with CXCR3 to define a pTFH subset that promotes IgG and IgA production [14], and PD-1. CCR7highCXCR5highCCR6high cells represented 1.2+/−0.9% of total CD4 T cells and a minority of these cells were PD-1high.

To analyze the ability of these populations to promote B cell differentiation, naïve and CM CD4 T cells from HIV-uninfected individuals were sorted based on expression of CCR7, CXCR5, CD150, CCR6 and PD-1 (Figure 1A), and cultured with autologous naïve B cells (CD19+CD27−IgD+) as previously described [14], [18] in the presence of staphylococcal enterotoxin B (SEB). Notably, our sorted naïve B cell population did not express isotype-switched immunoglobulin (Figure S1A) and culture conditions that lacked SEB did not induce immunoglobulin production (data not shown). Naïve and CM CCR7low CD4 T cells failed to promote B cell differentiation and immunoglobulin production whereas CM CCR7highCXCR5low cells induced limited production of IgM, IgG1 and IgG3 compared to the CCR7highCXCR5high populations (Figure 1B). The CCR7highCXCR5highCCR6highPD-1high population induced the greatest production of IgG1, IgG3 and IgA compared to the CXCR5low population. Prior studies defined pTFH cells based on surface expression of CXCR5, CCR6 and the lack of CXCR3 expression [14]. We found that the greatest help for immunoglobulin production was from CXCR5highCCR6high cell populations and, within those, from the PD-1high cells. We did not eliminate a small population of CXCR3+ cells in order to avoid removing a larger population of CXCR5highCCR6high cells that induce B cell differentiation (Figure S1B).
The cytokine profile of pTFH populations shared characteristics with other Thelper subsets, including TH1, TH17 and Treg cells. Supernatant from the CXCR5highCCR6highPD-1low coculture contained the greatest quantities of TNF-α, IL-2, and IL-17 compared to the CXCR5highCCR6lowPD-1high coculture (Figure 1C). Notably, the CXCR5highCCR6highPD-1high population, which promoted the greatest production of IgG1, IgG3 and IgA, showed the greatest IL-21 production, although at low levels.
Overall, CXCR5high CD4 T cell populations induced B cell immunoglobulin production, although the CXCR5highCCR6highPD-1high population did so most efficiently. However, this population is not characteristic of a TFH population found in secondary lymphoid organs, as coculture supernatants included a broad array of cytokines characteristic of TFH cells and multiple other Thelper subsets.
Progressive loss of pTFH cells in HIV infection
To determine the impact of HIV on pTFH populations, we compared pTFH cells from HIV-uninfected subjects and treatment-naïve HIV-infected subjects (Table S1) as a frequency of total CD4 cells. Irrespective of how pTFH cells were defined, there was a significant decrease in the pTFH population from HIV-infected subjects compared to HIV-uninfected subjects (Figure 2A). Subjects with CD4 counts greater than 200 had significantly lower pTFH populations, while subjects with CD4 counts less than 200 had the lowest frequency of all phenotypically defined pTFH populations. However, when we defined the CCR6highPD-1high population as a subset of the CXCR5high population, the frequency of the CCR6highPD-1high population increased in subjects with CD4 counts less than 200 (Figure S2A). The increase in PD-1high cells was likely due to immune activation in HIV infection, as we observed increases in the frequency of both PD-1high and ICOShigh cells within the CXCR5high population, with the greatest increases seen in samples with CD4 counts less than 200 (Figure S2A). We also observed a positive trend between CXCR5highPD-1high cells and serum concentrations of soluble CD14. (Figure S2A). For 10 HIV-infected individuals on whom we had longitudinal samples, we observed a loss of pTFH populations as a frequency of total CD4 T cells over 36 to 48 months (Figure 2B). However, the frequency of PD-1high, ICOShigh and CCR6highPD-1high cells within the CXCR5high population remained stable (Figure S2B).
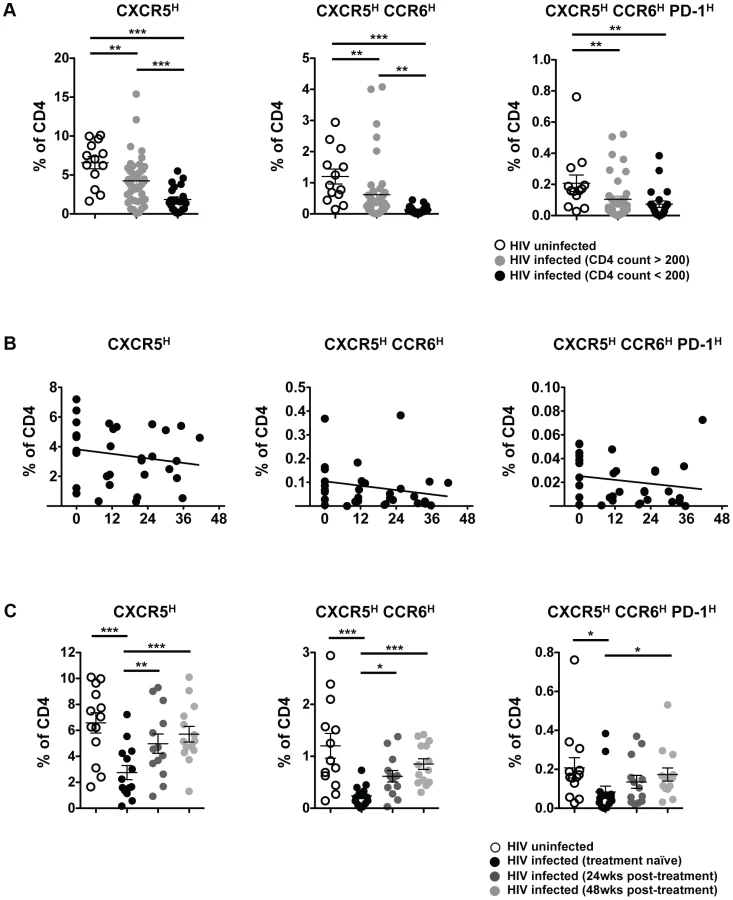
Next, we investigated the impact of ART on the frequency of pTFH within total CD4 T cells. Longitudinal analysis on samples from before and after 24 and 48 weeks of ART revealed a recovery of pTFH populations (Figure 2C). However, the frequency of PD-1high, ICOShigh and CCR6highPD-1high cells remained stable within the CXCR5high population (Figure S2C). Overall, HIV infection causes a loss of pTFH cells and ART promotes the recovery of these populations.
Impaired B cell help by pTFH cells in HIV infection
To investigate the impact of HIV on the ability of pTFH cells to support B cell differentiation, we performed co-culture experiments with pTFH cells from HIV-infected subjects. We focused on the CXCR5highCCR6high population that included both PD-1high and PD-1low cells due to limited cell numbers in HIV-infected subjects. Similar to previous results, the CXCR5highCCR6high population from HIV-uninfected subjects supported significantly more immunoglobulin production compared to the CXCR5low population. (Figure 3A). However, for HIV-infected subjects we observed less overall immunoglobulin production when CXCR5highCCR6high CD4 T cells were co-cultivated with naïve B cells. Furthermore, in viremic subjects, we observed increased IgM and IgG1 production in co-culture supernatants from the CXCR5low population, compared to HIV-uninfected subjects. Similar to HIV-uninfected subjects, we found that pTFH cells from HIV-infected subjects produced a broad spectrum of cytokines (Figure S3A).

Our data raise the possibility that some pTFH cells exhibit a CXCR5low phenotype in HIV infection. This phenotype could be due the down regulation of CXCR5 on pTFH cells, or indicate the existence of a unique CXCR5low pTFH population in chronic HIV infection. In order to distinguish these two possibilities, we investigated whether CXCL-13 impacts CXCR5 expression on CD4 T cells. We found that incubation of HIV-uninfected PBMC with CXCL-13 led to a decrease in frequency of CXCR5-positive CD4 T cells, presumably due to the internalization of CXCR5 (Figure 3B). Furthermore, in HIV infection we found that viral load positively correlated with CXCL-13 levels and negatively correlated with the frequency of CXCR5-positive CD4 T cells (Figure 3C). However, we did not observe a direct correlation between CXCL13 levels and the frequency of CXCR5-positive CD4 T cells. Importantly, we also found that in vitro infection of CXCR5-expressing CD4 T cells did not impact CXCR5 surface expression (Figure S3B). Therefore, our data support the possibility that in untreated HIV infected individuals, increased levels of CXCL-13 could effect CXCR5 surface expression on pTFH cells.
Defective cytokine production of pTFH in HIV infection
TFH-dependent B cell differentiation requires IL-21. To characterize directly cytokine production from pTFH cells from HIV-uninfected and HIV-infected subjects, we performed intracellular cytokine staining after ex vivo SEB stimulation. In addition to surface markers used to define pTFH cells, we detected CD154, IFN-γ, IL-2, IL-17 and IL-21 (Figure 4A). In HIV-uninfected individuals, a minority of CD154-positive, cytokine-positive cells express a CCR7high phenotype (10.1% of IFN-γ positive cells; 28% of IL-2-positive cells; 19.4% of IL-17-positive cells and 17.9% of IL-21-positive cells), while a gradual reduction of cytokine production was found in further differentiated cells based on CXCR5 and CCR6 expression (Figure 4B). However, for all of the cytokines detected, we observed a population of cells that were CCR7highCXCR5highCCR6high, including IL-21-producing cells. Overall, we determined that a mean of 4.5% of CD154-positive IL-21-positive cells were CCR7highCXCR5highCCR6high (Figure 4B). However, this pTFH population also produced IFN-γ, IL-2 and IL-17 (0.8% of IFN-γ positive cells; 9.0% of IL-2-positive cells and 7.1% of IL-17-positive cells).

Next, we analyzed cytokine production from HIV-infected subjects off-treatment. Overall, we observed a loss of cytokine-producing cells from the CCR7high population and a general shift towards the CXCR5lowCCR6low population (Figure 4A). Thus, we observed a loss of CCR7highCXCR5highCCR6high pTFH cells that produce IL-2, IL-17 and IL-21 (Figure 3B; IL-2 : 9.0% for HIV-negative vs 2.0% for HIV-positive; IL-17 : 7.1% for HIV-negative vs 2.2% for HIV-positive; IL-21 : 4.5% for HIV-negative vs 1.1% for HIV-positive).
To analyze HIV-specific cells, PBMC were stimulated with Gag peptide pools and analyzed for cytokine expression. Very few IL-2-positive and IL-17-positive cells were detected within the CM compartment (data not shown). Gag-specific IFN-γ and IL-21-producing cells were detected, however, compared to SEB-stimulation fewer HIV-specific cells expressed CCR7 (4.4% vs 10.7% of IFN-γ positive cells; 3.5% vs 11.9% of IL-21-positive cells for Gag and SEB stimulation, respectively). A majority of HIV-specific cells were not CCR7highCXCR5highCCR6high (Figure 4C; 0.4% of IFN-γ positive cells and 0.9% of IL-21-positive cells were CCR7highCXCR5highCCR6high).
Overall, we observed IL-21 production from the CCR7highCXCR5highCCR6high pTFH population, although we detected the most IL-21 in non-pTFH cells, which were CCR7low and CXCR5low. In addition to IL-21, the CCR7highCXCR5highCCR6high pTFH population produced IL-2 and IL-17, cytokines characteristic of TH1 and TH17 cells, respectively. However, from HIV-infected individuals we observed a loss of CCR7highCXCR5highCCR6high cells making IL-2, IL-17 and IL-21.
No relationship between pTFH cells and neutralization activity
Previous studies have described a relationship between the frequency of peripheral CXCR5high cells and memory B cells and antibody titers with vaccination [16]. Therefore, we analyzed the relationship between the frequency of pTFH cells and IgG-positive memory B cells in PBMC from HIV-infected individuals. We found no significant correlation between the frequency of pTFH cells and IgG-positive B cells (Figure 5A). Similarly, we failed to detect a relationship between the frequency of pTFH and HIV-1 Env-specific antibody titers or total plasma IgG levels (data not shown).

It has also been reported that PD-1high CD4 T cells in blood are associated with cross-clade neutralizing antibody responses during HIV infection [19] and these PD-1high CD4 T cells may represent a population of pTFH cells. Thus, the relationship between pTFH cells and neutralization activity was analyzed using HIV-infected samples classified as good neutralizers (median ID50>100) or poor neutralizers (median ID50<100) [20]. Irrespective of how pTFH cells were defined, we failed to find any relationship between neutralization activity and pTFH cells (Figure 5B).
Relationship between pTFH cells and TFH cells in human tonsil
While pTFH cells induce B cell differentiation and immunoglobulin secretion in vitro, the relationship between pTFH and TFH cells in secondary lymphoid organs remains unclear. Our in vitro coculture studies indicated the greatest isotype-switched immunoglobulin production was elicited from B cells cocultivated with CXCR5highCCR6high pTFH cells (Figure 1B). Therefore, we investigated the expression of CCR6 on TFH (CXCR5highPD-1high) and non-TFH (CXCR5lowPD-1low) tonsil cells to determine if the CXCR5highCCR6high pTFH population is related to TFH cells within secondary lymphoid organs (Figure 6A). The lowest frequency of CCR6high cells was found within the CXCR5highPD-1high compartment (1.5% of CXCR5highPD-1high cells) and the greatest frequency of CCR6high cells within the non-TFH compartment (9% of CXCR5lowPD-1low cells; Figure 6B).

Similarly, RNA sequence data from the CXCR5highCCR6highPD-1high pTFH population more closely resembles a memory, non-TFH CD4 T cell population from the tonsil (CM CD57lowPD-1dimCCR7highCCR5lowCXCR4low) as compared to the non-germinal center TFH population (CM CD57lowPD-1highCCR7lowCXCR5high) or the GC-TFH population (CM PD-1highCD57high; Figure 6C). In agreement with previous reports [5], [17], tonsil TFH populations expressed higher levels of BCL6, IL-21, and CXCL13, and lower levels of PRDM1 and S1PR1 compared to the non-TFH memory population. The pTFH population from HIV-uninfected individuals expressed comparable levels of S1PR1 and PRDM1 to the non-TFH memory population in the tonsil (Figure 6). We also observed lower transcript levels of MAF, BCL6, IL-21, and CXCL-13 in the pTFH population compared to tonsillar TFH populations. Importantly, MAF protein expression was highest in the CCR6highPD-1high pTFH population compared to other peripheral populations, although still lower than tonsillar TFH cells. (Figure 6D). For many of the selected genes, pTFH cells from HIV-infected subjects were comparable to pTFH from HIV-uninfected individuals, however, we observed greater transcript levels of activation molecules such as ICOS and CD69. Additionally, the levels of IL-21 were decreased in pTFH cells from HIV-infected individuals, supporting earlier results (Figure 4B). Collectively, our data suggest the pTFH population characterized as CXCR5highCCR6high most closely resembles a non-TFH memory population in the tonsil.
Discussion
The development and nature of human TFH memory cells following an effector immune response are not known. The ability to define a population of memory TFH cells in PBMC (pTFH) would help inform our understanding of CD4 T cell dynamics within lymphoid tissue during vaccination or infection. Studies of chronic infection may be helpful in this regard [21]. Whether the accumulation of TFH cells during chronic infection [4], [15] impacts the TFH memory population is of particular interest, especially if memory TFH cells migrate between lymphoid organs and peripheral tissues. Recent studies [14], [16] have suggested that circulating CXCR5high CD4 T cells may represent the peripheral counterparts of TFH cells. However, the relationship between pTFH and TFH cells within secondary lymphoid organs remains unclear. Therefore, it is of great relevance to determine if pTFH cells originate from GC-TFH cells and represent a memory TFH population, reflect a precursor population that differentiates into GC-TFH upon re-exposure to antigen, or both. Our studies begin to address these issues by further defining pTFH cells, comparing pTFH cells to tonsillar TFH cells, and analyzing the effect of HIV on these cells.
In concordance with previous studies, we showed that circulating CXCR5high CD4 T cells support B cell differentiation in vitro [14], [17]. A majority of the CXCR5high cells expressed CD150, and while CD150 was used for gating in the co-culture assays, we found it did not impact the loss of pTFH cells or effect our results with respect to loss of pTFH cells, recovery with ART or lack of association with B cell or antibody responses (data not shown). However, within the CXCR5high population the expression of CCR6 and PD-1 did further define pTFH populations with differential abilities for naïve B cell help and isotype switching. Thus, pTFH cell populations support both the activation and maturation of naïve B cells, and immunoglobulin isotype switching. Correspondingly, the individual pTFH populations produced cytokines associated with B cell maturation and survival, such as IL-21 [22], IL-2 [23] and IL-17 [24], in contrast to TFH cells within secondary lymphoid tissue, which display a limited cytokine profile that includes IL-4, IL-10 and IL-21, but compromised production of IL-2 and IL-17 [4]. Whether these pTFH populations represent different stages of TFH memory development or originate from separate CD4 T cell populations within lymphoid tissue [25] is still unclear.
In order to better understand the relationship between TFH and pTFH cells, we compared gene expression levels between pTFH and tonsillar CD4 T cell populations and focused on genes important for TFH differentiation, migration, and function. We found that the pTFH population with the greatest B cell helper function most closely resembled a CM, non-TFH CD4 T cell subset within the tonsil. While our studies do not directly address the relationship between GC-TFH in lymph nodes and circulating CD4 T cells from the same patients, our data challenge whether pTFH are memory TFH cells. A recent study reported that germinal center TFH cells in mice migrate throughout the follicle, but generally do not leave the follicle to enter the blood [26]. While it is conceivable that pTFH cells represent a very minor population of TFH cells that exit the follicle, it is also possible that pTFH cells are reflective of a precursor TFH population that exits the lymphoid organ and enters the circulation before entering the follicle. However, while we find the CXCR5highCCR6highPD-1high pTFH population does not resemble a memory TFH population, Locci and colleagues found a CXCR5+CXCR3-PD-1+ pTFH subset that functionally and transcriptionally resembles a memory TFH population [27]. A recent study in mice reported that memory TFH cells have reduced mRNA expression of TFH markers such as Bcl6, IL-21, ICOS and PD-1 compared to the effector TFH population [28], indicating the expression of these molecules may change depending on the phase of infection. Therefore, further investigation of pTFH subsets and their relationship to memory and effector populations at multiple stages of infection is needed.
pTFH and naïve B cell co-cultures from HIV-infected subjects produced fewer immunoglobulins compared to co-cultures from HIV-uninfected subjects. The observed defect in immunoglobulin production is likely due to impaired pTFH help to B cells instead of B cell dysfunction, as co-cultures included naïve B cells rather than memory B cells that exhibit abnormalities in HIV infection [29]. Furthermore, while co-culture supernatants from HIV-infected subjects demonstrated a heterogeneous cytokine profile, similar to HIV-uninfected subjects, intracellular cytokine staining showed that fewer CCR7highCXCR5highCCR6high pTFH cells produced IL-2, IL-17 and IL-21 in chronic HIV infection compared to HIV-uninfected individuals. Furthermore, gene expression analysis of HIV-infected pTFH revealed fewer IL-21 and IL-4 transcripts, although the overall levels of cytokine transcripts were low.
Recent studies have shown TFH cells within secondary lymphoid organs accumulate in some donors or animals during chronic HIV/SIV infection and that TFH accumulation is associated with GC B cell expansion and increased serum immunoglobulin concentrations [4], [22], [30]. In contrast to TFH cells, our studies revealed pTFH cells consistently decrease in chronic HIV infection, with disease progression resulting in a greater reduction of these compartments within the total CD4 T cell population. However, it should be noted that we were unable to analyze TFH cells within secondary lymphoid organs from these subjects and therefore we are unable to directly compare the frequency of pTFH cells and TFH cells from the same individual. The differences between the increase in TFH cells and decrease in pTFH cells may be due to differences in disease state (i.e. early vs late infection) or represent a steady state of TFH cells trafficking between the lymphoid tissue and the blood. The decreased frequency of pTFH in the blood may indicate impaired ability of TFH to exit the lymph node in chronic HIV infection where the tissue architecture is not intact. Alternatively, the decreased frequency of pTFH in the blood may be a result of pTFH trafficking to secondary lymphoid organs. In agreement with previous studies [14], [17], we found a majority of CXCR5high cells express CCR7, and it has previously been suggested that pTFH cells migrate to secondary lymphoid organs upon infection due their expression of CCR7 and CD62L [14].
A confounding factor with regard to how we interpret the decrease in pTFH cells is that we also found a reduction in the surface expression of CXCR5 on CD4 T cells in chronic HIV infection, which may result from increased sera levels of CXCL-13 [31], [32]. Furthermore, our co-culture data indicate that CXCR5low CD4 T cells from viremic subjects can induce some B cell differentiation. These data support the possibility that in chronic HIV infection, a subset of functional pTFH cells may be phenotypically defined as CXCR5low. Additionally, it should be noted that analysis of cellular subsets within the CXCR5high population in chronic HIV infection revealed the frequency of CCR6highPD-1high cells increased. These results are consistent with a state of generalized immune activation, as we also observed increased surface expression of ICOS on CXCR5high and CXCR5highPD-1high cells, and a positive association between the frequency of PD-1high cells within the CXCR5high population and serum concentrations of soluble CD14 [33]. Similarly, gene expression analysis indicated increased transcript levels of activation markers, such as ICOS and CD69 within the pTFH population during HIV infection. Overall, these data emphasize the difficulty in defining pTFH cells in chronic HIV infection and understanding the relationship between pTFH cells and TFH cells.
The uncertain definition of pTFH cells in HIV infection may provide an explanation as to why we were unable to identify correlations between pTFH populations and circulating IgG-positive memory B cells, or between pTFH cells and HIV-specific IgG (data not shown). Furthermore, we found no correlation between the frequency of pTFH and the neutralization activity of a well-characterized cohort of HIV-infected donors [20]. However, the absence of a correlation between pTFH cells and circulating HIV Env-specific IgG may also be explained by the lack of a time-dependent association (early vs. late infection) between TFH and pTFH cells, or indicate that the generation of IgG and broadly neutralizing antibodies is regulated by parameters other than pTFH, confounded by T-cell independent antibody production commonly observed in HIV infection [34] or generalized immune activation. Thus, our data challenge the application of the pTFH population as a surrogate of GC TFH-B cell interactions in chronic HIV infection. While our studies did not find a correlation between pTFH cells and neutralizing antibodies, several recent studies, each with a different definition of pTFH cells, have reported an association with antibody responses during vaccination, infection or autoimmune disease [27], [35]–[37]. Therefore, further studies are needed to establish the association between pTFH subsets and the generation of neutralizing antibodies, especially in HIV infection.
Overall, our data indicate that a range of circulating CD4 T cell populations can provide B cell help, possibly through differential secretion of soluble factors and/or cell-cell contact interactions [17], [35] and that HIV infection results in loss of these cells over time, but with relative increases within the CXCR5high compartment which may be explained by immune activation. Furthermore, we did not find any association between pTFH and measures of B cell function such as HIV neutralization breadth/potency, HIV-specific IgG, or total IgG, suggesting application of this population as a surrogate of GC TFH-B cell interactions during HIV infection may be limited. A better understanding of the differentiation process and the developmental relationship between pTFH subsets and lymph node TFH cells is critical for the establishment of reliable peripheral blood CD4 T cell correlates for monitoring infection - or vaccine-associated B cell responses.
Materials and Methods
Ethics statement
Signed informed consent was obtained in accordance with the Declaration of Helsinki and approved by the appropriate Institutional Review Board. Tonsil cells were acquired from anonymized discarded pathologic specimens from Children's National Medical Center (CNMC) under the auspices of the Basic Science Core of the District of Columbia Developmental Center for AIDS Research. The CNMC Institutional Review Board determined that study of anonymized discarded tonsils did not constitute ‘human subjects research.’
Subjects
Fresh HIV-uninfected peripheral blood mononuclear cells (PBMC) were obtained from individuals participating in the NIH research apheresis program. Fresh HIV-infected blood was obtained from the Vaccine Research Center Clinic or Drexel University College of Medicine. Frozen HIV-infected PBMC were obtained from three study populations (Table S1). For untreated HIV infection, cells were obtained from volunteers who participated in a therapeutic vaccination trial (no efficacy was observed) conducted in the 1990's prior to the advent of combination antiretroviral therapy (cART) [38]. The second study population consisted of donors from a cohort used to identify individuals with HIV broadly neutralizing antibodies [20]. To study the effect of cART, we obtained PBMC from HIV-infected donors participating in AIDS Clinical Trials Group study A5142 prior to initiation of cART and 24 and 48 weeks post-therapy [39], [40]. PBMC and tonsil cells were cultured in RPMI 1640 supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100 U/mL penicillin and 100 µg/mL streptomycin (Invitrogen).
Antibodies
Directly conjugated antibodies were acquired from the following: (1) BD Biosciences: CD3-H7APC, CXCR5-Alexa488 (RF8B2), CCR7-Alexa700, IgG-APC, IFN-γ-Alexa700 and IL-21-Alexa647 (3A3-N2.1) (2) Beckman Coulter: CD45RO-ECD and CD27-PC5 (3) Biolegend: CCR7-BV421, CCR6-PE (TG7/CCR6), CCR6-Alexa647 (TG7/CCR6), CD20-BV570, CD150-PE, IL-2-BV605, IL-17a-Cy5.5PerCP and CD154-Cy5PE (4) Invitrogen: CD4-Cy5.5PE, CD27-QD655, CD27-QD605 and CD19-PacBlue (5) Southern Biotech: IgD-FITC and IgD-PE (6) eBioscience: cmaf-eFluor660 (sym0F1), CXCR5-PerCP-efluor710 (MU5UBEE). Biotinylated anti-PD-1 was from R&D and streptavidin-Cy7PE (or QD655) was from Molecular Probes. The following antibodies were conjugated in our lab: CD19-QD705 and CD57-QD565. Quantum dots and Aqua amine viability dye were obtained from Invitrogen.
Polychromatic flow cytometry
Phenotypic analysis
1–2×106 PBMC were incubated with Aqua-dye and surface stained with titrated amounts of anti-CD3, anti-CD4, anti-CD27, anti-CD45RO, anti-CCR7, anti-CXCR5, anti-CD150, anti-CCR6, anti-PD-1 and anti-CD19. Post-wash, cells were incubated with fluorescent-conjugated streptavidin, washed and fixed with 1% paraformaldehyde.
Intracellular cytokine staining
3×106 PBMC were incubated in 1 mL of medium containing brefeldin A (10 ug/mL) in the absence or presence of HIV-1 Gag-peptide pools (15mers overlapping by 11 residues; NIH AIDS Research and Reference Reagent Program) or 1 ug/mL SEB (Sigma) for 6 hours. Cells were surface stained, permeabilized (Cytofix/Cytoperm kit; BD Biosciences), and stained with anti-CD3, anti-IFN-γ, anti-IL-2, anti-IL-17a, anti-IL-21 and anti-CD154. Events were collected on a modified LSRII flow cytometer (BD Immunocytometry Systems) and electronic compensation was performed with antibody capture beads (BD Biosciences). Data were analyzed using FlowJo Version 9.6 (TreeStar).
T and B cell culture
Co-culture experiments were performed with freshly isolated PBMC. 5×104 CD4 T cell populations were sorted based on expression of CCR7, CXCR5, CD150, CCR6 and PD-1 and cultured with 5×104 autologous naïve B cells (1∶1 ratio) in the presence of SEB (0.5 µg/ml). Supernatants harvested on Day 2 were analyzed for cytokines using Luminex technology (Milliplex MAP Kit, HTH17MAG-14K, Millipore). The lower limit of detection (LOD) was set at the lowest concentration on the standard curve and values below the LOD were counted as zero. Supernatants collected on Day 12 were analyzed for immunoglobulins (Milliplex MAP Kit, HGAMMAG-301K). Some supernatants exceeded the saturation limit of the standard curves for IgM and IgG3. These values were included in the analysis and quantified as being equivalent to the highest determined concentration.
ELISA
Soluble CD14 and CXCL-13 (R&D Systems) were measured in plasma or sera from HIV-infection patients according to the manufacturer's instructions.
CXCL-13 treatment
Freshly isolated PBMCs were incubated with recombinant human CXCL-13 (R&D Systems) at 37°C or 4°C and analyzed for CXCR5 surface expression by FACS.
Illumina deep sequencing of messenger RNA
CD4 T cell populations were sorted from uninfected PBMC (n = 5), HIV-infected PBMC (n = 5) and uninfected human tonsils (n = 4) based on expression of CCR7, CXCR5, CD150, CCR6 and PD-1 for PBMC and CD57, PD-1, CCR7, CXCR5, CCR5 and CXCR4 for tonsils. Total RNA was purified from sorted cell populations and treated with DNAse I (Ambion) to minimize genomic DNA contamination. Polyadenylated RNA was isolated using Oligo-dT Dynabeads (Life Technologies), chemically fragmented, and used to construct barcoded Illumina Truseq libraries. Libraries were size-selected, quantified, pooled, size-selected and quantified again, and clustered on an Illumina Truseq Paired-End Flowcell v3. The flowcell was sequenced on an Illumina HiSeq 2000 in a 2×75-base paired-end, indexed run. Adaptor sequence was trimmed from the raw sequencing reads using Trimmomatic. The trimmed sequencing reads were subsequently aligned to the human genome (hg19) using TopHat 2. Differential expression testing was done using Cufflinks 2 and visualization of differential expression was done using the R package cummerbund. Accession numbers of the selected genes are shown in Supporting Table S2.
Virus neutralization
Neutralization activity of patient sera was determined against 20 viral isolates using a TZM-bl neutralization assay as previously described [20].
In vitro infection
Freshly isolated PBMCs were stimulated with PHA (10 µg/ml). After 12 hours stimulation, CXCR5high cells were sorted by FACS Aria based on surface molecule expression and infected by a multiplicity of infection (MOI) of 0.01 with either HIV NL-E or HIV NLAD8-E [41]. The infected cells were cultured in the presence of 50 U/ml recombinant human interleukin-2 (R&D) for 5 days and analyzed for CXCR5 expression by FACS.
Statistics
Experimental variables were analyzed using the nonparametric Mann-Whitney U test, the Wilcoxon matched-pairs signed rank test or the Friedman test with Dunn's multiple comparison post-test. Correlation analysis was performed using the nonparametric Spearman test. Error bars depict mean+SEM in all bar graphs shown. The GraphPad Prism statistical analysis program (GraphPad Software, version 5.0) was used throughout.
Supporting Information
Zdroje
1. CrottyS (2011) Follicular helper CD4 T cells (TFH). Annu Rev Immunol 29 : 621–663.
2. HaynesNM, AllenCD, LesleyR, AnselKM, KilleenN, et al. (2007) Role of CXCR5 and CCR7 in follicular Th cell positioning and appearance of a programmed cell death gene-1high germinal center-associated subpopulation. J Immunol 179 : 5099–5108.
3. MaCS, SuryaniS, AveryDT, ChanA, NananR, et al. (2009) Early commitment of naive human CD4(+) T cells to the T follicular helper (T(FH)) cell lineage is induced by IL-12. Immunol Cell Biol 87 : 590–600.
4. PetrovasC, YamamotoT, GernerMY, BoswellKL, WlokaK, et al. (2012) CD4 T follicular helper cell dynamics during SIV infection. J Clin Invest 122 : 3281–3294.
5. KimCH, LimHW, KimJR, RottL, HillsamerP, et al. (2004) Unique gene expression program of human germinal center T helper cells. Blood 104 : 1952–1960.
6. YuD, VinuesaCG (2010) The elusive identity of T follicular helper cells. Trends Immunol 31 : 377–383.
7. FaheyLM, WilsonEB, ElsaesserH, FistonichCD, McGavernDB, et al. (2011) Viral persistence redirects CD4 T cell differentiation toward T follicular helper cells. J Exp Med 208 : 987–999.
8. Glatman ZaretskyA, TaylorJJ, KingIL, MarshallFA, MohrsM, et al. (2009) T follicular helper cells differentiate from Th2 cells in response to helminth antigens. J Exp Med 206 : 991–999.
9. NurievaRI, ChungY, HwangD, YangXO, KangHS, et al. (2008) Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity 29 : 138–149.
10. NurievaRI, ChungY, MartinezGJ, YangXO, TanakaS, et al. (2009) Bcl6 mediates the development of T follicular helper cells. Science 325 : 1001–1005.
11. YuD, RaoS, TsaiLM, LeeSK, HeY, et al. (2009) The transcriptional repressor Bcl-6 directs T follicular helper cell lineage commitment. Immunity 31 : 457–468.
12. LuthjeK, KalliesA, ShimohakamadaY, BelzGT, LightA, et al. (2012) The development and fate of follicular helper T cells defined by an IL-21 reporter mouse. Nat Immunol 13 : 491–498.
13. PepperM, JenkinsMK (2011) Origins of CD4(+) effector and central memory T cells. Nat Immunol 12 : 467–471.
14. MoritaR, SchmittN, BentebibelSE, RanganathanR, BourderyL, et al. (2011) Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 34 : 108–121.
15. LindqvistM, van LunzenJ, SoghoianDZ, KuhlBD, RanasingheS, et al. (2012) Expansion of HIV-specific T follicular helper cells in chronic HIV infection. J Clin Invest 122 : 3271–3280.
16. PallikkuthS, ParmigianiA, SilvaSY, GeorgeVK, FischlM, et al. (2012) Impaired peripheral blood T-follicular helper cell function in HIV-infected nonresponders to the 2009 H1N1/09 vaccine. Blood 120 : 985–993.
17. ChevalierN, JarrossayD, HoE, AveryDT, MaCS, et al. (2011) CXCR5 expressing human central memory CD4 T cells and their relevance for humoral immune responses. J Immunol 186 : 5556–5568.
18. BreitfeldD, OhlL, KremmerE, EllwartJ, SallustoF, et al. (2000) Follicular B helper T cells express CXC chemokine receptor 5, localize to B cell follicles, and support immunoglobulin production. J Exp Med 192 : 1545–1552.
19. MikellI, SatherDN, KalamsSA, AltfeldM, AlterG, et al. (2011) Characteristics of the earliest cross-neutralizing antibody response to HIV-1. PLoS Pathog 7: e1001251.
20. Doria-RoseNA, KleinRM, DanielsMG, O'DellS, NasonM, et al. (2010) Breadth of human immunodeficiency virus-specific neutralizing activity in sera: clustering analysis and association with clinical variables. J Virol 84 : 1631–1636.
21. HaleJS, YoungbloodB, LatnerDR, MohammedAU, YeL, et al. (2013) Distinct Memory CD4(+) T Cells with Commitment to T Follicular Helper - and T Helper 1-Cell Lineages Are Generated after Acute Viral Infection. Immunity 38 : 805–817.
22. LintermanMA, BeatonL, YuD, RamiscalRR, SrivastavaM, et al. (2010) IL-21 acts directly on B cells to regulate Bcl-6 expression and germinal center responses. J Exp Med 207 : 353–363.
23. Le GallouS, CaronG, DelaloyC, RossilleD, TarteK, et al. (2012) IL-2 requirement for human plasma cell generation: coupling differentiation and proliferation by enhancing MAPK-ERK signaling. J Immunol 189 : 161–173.
24. DoreauA, BelotA, BastidJ, RicheB, Trescol-BiemontMC, et al. (2009) Interleukin 17 acts in synergy with B cell-activating factor to influence B cell biology and the pathophysiology of systemic lupus erythematosus. Nat Immunol 10 : 778–785.
25. BentebibelSE, SchmittN, BanchereauJ, UenoH (2011) Human tonsil B-cell lymphoma 6 (BCL6)-expressing CD4+ T-cell subset specialized for B-cell help outside germinal centers. Proc Natl Acad Sci U S A 108: E488–497.
26. ShulmanZ, GitlinAD, TargS, JankovicM, PasqualG, et al. (2013) T follicular helper cell dynamics in germinal centers. Science 341 : 673–677.
27. LocciM, Havenar-DaughtonC, LandaisE, WuJ, KroenkeMA, et al. (2013) Human Circulating PD-1CXCR3CXCR5 Memory Tfh Cells Are Highly Functional and Correlate with Broadly Neutralizing HIV Antibody Responses. Immunity 39 : 758–769.
28. HaleJS, YoungbloodB, LatnerDR, MohammedAU, YeL, et al. (2013) Distinct memory CD4+ T cells with commitment to T follicular helper - and T helper 1-cell lineages are generated after acute viral infection. Immunity 38 : 805–817.
29. MalaspinaA, MoirS, DiPotoAC, HoJ, WangW, et al. (2008) CpG oligonucleotides enhance proliferative and effector responses of B Cells in HIV-infected individuals. J Immunol 181 : 1199–1206.
30. HarkerJA, LewisGM, MackL, ZunigaEI (2011) Late interleukin-6 escalates T follicular helper cell responses and controls a chronic viral infection. Science 334 : 825–829.
31. CagigiA, MowafiF, Phuong DangLV, Tenner-RaczK, AtlasA, et al. (2008) Altered expression of the receptor-ligand pair CXCR5/CXCL13 in B cells during chronic HIV-1 infection. Blood 112 : 4401–4410.
32. WidneyDP, BreenEC, BoscardinWJ, KitchenSG, AlcantarJM, et al. (2005) Serum levels of the homeostatic B cell chemokine, CXCL13, are elevated during HIV infection. J Interferon Cytokine Res 25 : 702–706.
33. SandlerNG, WandH, RoqueA, LawM, NasonMC, et al. (2011) Plasma levels of soluble CD14 independently predict mortality in HIV infection. J Infect Dis 203 : 780–790.
34. HeB, QiaoX, KlassePJ, ChiuA, ChadburnA, et al. (2006) HIV-1 envelope triggers polyclonal Ig class switch recombination through a CD40-independent mechanism involving BAFF and C-type lectin receptors. J Immunol 176 : 3931–3941.
35. BentebibelSE, LopezS, ObermoserG, SchmittN, MuellerC, et al. (2013) Induction of ICOS+CXCR3+CXCR5+ TH Cells Correlates with Antibody Responses to Influenza Vaccination. Sci Transl Med 5 : 176ra132.
36. SpensieriF, BorgogniE, ZeddaL, BardelliM, BuricchiF, et al. (2013) Human circulating influenza-CD4+ ICOS1+IL-21+ T cells expand after vaccination, exert helper function, and predict antibody responses. Proc Natl Acad Sci U S A 110 : 14330–14335.
37. HeJ, TsaiLM, LeongYA, HuX, MaCS, et al. (2013) Circulating Precursor CCR7loPD-1hi CXCR5+ CD4+ T Cells Indicate Tfh Cell Activity and Promote Antibody Responses upon Antigen Reexposure. Immunity 39 : 770–781.
38. BirxDL, Loomis-PriceLD, AronsonN, BrundageJ, DavisC, et al. (2000) Efficacy testing of recombinant human immunodeficiency virus (HIV) gp160 as a therapeutic vaccine in early-stage HIV-1-infected volunteers. rgp160 Phase II Vaccine Investigators. J Infect Dis 181 : 881–889.
39. HaubrichRH, RiddlerSA, DiRienzoAG, KomarowL, PowderlyWG, et al. (2009) Metabolic outcomes in a randomized trial of nucleoside, nonnucleoside and protease inhibitor-sparing regimens for initial HIV treatment. AIDS 23 : 1109–1118.
40. RiddlerSA, HaubrichR, DiRienzoAG, PeeplesL, PowderlyWG, et al. (2008) Class-sparing regimens for initial treatment of HIV-1 infection. N Engl J Med 358 : 2095–2106.
41. CooperA, GarciaM, PetrovasC, YamamotoT, KoupRA, et al. (2013) HIV-1 causes CD4 cell death through DNA-dependent protein kinase during viral integration. Nature 498 : 376–379.
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2014 Číslo 1
- Farmakovigilanční studie perorálních antivirotik indikovaných v léčbě COVID-19
- Jak souvisí postcovidový syndrom s poškozením mozku?
- Měli bychom postcovidový syndrom léčit antidepresivy?
- 10 bodů k očkování proti COVID-19: stanovisko České společnosti alergologie a klinické imunologie ČLS JEP
Nejčtenější v tomto čísle
- Lyme Disease: Call for a “Manhattan Project” to Combat the Epidemic
- Origin, Migration Routes and Worldwide Population Genetic Structure of the Wheat Yellow Rust Pathogen f.sp.
- IFNγ/IL-10 Co-producing Cells Dominate the CD4 Response to Malaria in Highly Exposed Children
- Human and Plant Fungal Pathogens: The Role of Secondary Metabolites